Questa discussione è bloccata
Questa discussione è bloccata
APPUNTI SU UNA RACCOLTA AGARICOIDE
DI RUSSULA MESSAPICA
di
Paolo Picciola* Marino Zugna** Silva Refellato *** Bruno De Ruvo ****Paolo De Martin *****
(*) Paolo Picciola - Strada di Fiume, 52 - 34137 Trieste (TS) paolo.picciola2@gmail.com
(**) Marino Zugna - Località Riostorto 7, 34015 Muggia (TS) marinozugna@gmail.com
(***) Refellato Silva - Via Noalese 41/4, 30037 Scorzè (VE) refellatosilva@gmail.com
(****) Bruno de Ruvo - Viale E. Crucioli, 26, 64100 Teramo (TE) colorificio@centroparati.it
(****) De Martin Paolo - Via Dei Pellegrini 41/A, 34139 Trieste (TS) paolo.50p@libero.it
Introduzione
Gli autori descrivono una raccolta del portamento agaricoide di Russula messapica effettuata da uno di noi (B.de R.) durante il 79° Comitato scientifico Nazionale - Lecce - 7/11 novembre 2018.
Della raccolta si riportano le caratteristiche macro e microscopiche nonché le immagini a colori dei basidiomi fotografati in situ e dei principali caratteri microanatomici rilevati.
Sono riportate alcune osservazioni di ordine sistematico riguardanti questa rara specie che, in virtù delle sue peculiari caratteristiche macro e microscopiche, è stata inserita dallo specialista Sarnari nella Sezione Messapicae.
Vengono brevemente citati alcuni confronti con le russule velangiocarpiche europee contrassegnate da una spettacolare reazione rosso intenso agli alcali.
L’ultima parte è dedicata alla sua forma angiocarpica inizialmente descritta con il nome di Macowanites messapicoides e che recentemente è stata riconosciuta una semplice forma angiocarpica di Russula messapica.
Key words: Agaricomycetes, Russulales, Russula messapica, Puglia.
Materiali e metodi
La descrizione dei caratteri macroscopici è stata ricavata analizzando i basidiomi freschi fotografati in situ con l’ausilio di una fotocamera Nikon Df, obiettivo Nikkor 60 mm f/2.8.
Lo studio microscopico è stato condotto su esemplari essiccati preventivamente idratati con l’idrossido di potassio (KOH) al 2%.
I preparati sono stati osservati in acqua e dove necessario si è adoperato il Rosso Congo Ammoniacale al 2% per evidenziare le differenti tipologie cellulari. Le misure microscopiche sono state eseguite con il software di calcolo Mycométre gentilmente fornitoci dal sig. Georges Fannechère e prelevabile all’indirizzo http://mycolim.free.fr.
Le misure sporali si riferiscono a 100 unità.
Per le rimanenti tipologie cellulari, dove possibile, si sono eseguite un minimo di venti misure a tipologia.
Le misure sottolineate di tutti gli elementi microscopici rappresentano i valori medi.
Le foto concernenti la microscopia, sono state eseguite con l’ausilio di una fotocamera Reflex EOS 50D, posta sul terzo occhio di un microscopio biologico Optech Biostar B5, supportato da ottiche Plan-APO, illuminazione alogena 12V-50 W a luce riflessa, con regolatore d'intensità.
Per quanto riguarda la nomenclatura adottata si è fatto riferimento a Index Fungorum http://www.indexfungorum.org/ e MykoBank http://www.mycobank.org.
La collezione d’erbario è conservata nell'erbario del Gruppo di Muggia e del Carso della A.M.B. con il numero identificativo 7712.
TASSONOMIA
Russula messapica Sarnari ex Sarnari
Bollettino dell'Associazione Micologica ed Ecologica Romana 18: 12 (1990)
Sinonimi
Russula messapicoides (Llistos. & J.M. Vidal) Trappe & T.F. Elliott, Fungal Systematics and Evolution 1: 236 (2018).
Macowanites messapicoides Llistos. & J.M. Vidal, Rivista di Micologia 38 (2): 155 (1995).
Russula messapica var. messapicoides (Llistos. & J.M. Vidal) J.M. Vidal, Llistos., Kaounas & P. Alvarado, Persoonia 42: 167 (2019).
Sistematica: Fungi, Dikarya, Basidiomycota, Agaricomycotina, Agaricomycetes, Russulales, Russulaceae, Russula.
Descrizione della raccolta
Pileo fino a 50 mm di diametro, poco carnoso e piuttosto fragile, rapidamente espanso, leggermente depresso, con il margine un po' irregolare, flessuoso, ottuso, solo appena scanalato a maturità, cuticola asportabile per un ampio tratto, liscia e brillante con l’umido, presto asciutta e opaca, di colore giallo tuorlo, con tinte aranciate e arancio-rossastre più evidenti al disco.
Lamelle uguali, larghe e ventricose, sottili, ottuse in avanti, con il filo intero e concolore, marcatamente intervenate sul fondo, fragili, gialline a maturità, sfumate di aranciato.
Stipite cilindrico, piuttosto regolare, pieno, poi lacunoso, asciutto e finemente corrugato per tutta la lunghezza, dapprima bianco, un po' ingrigente con l’umidità, si tinge progressivamente di giallo cadmio a partire dalla base.
Carne molle e piuttosto delicata soggetto alla putrefazione, bianca, giallastra nelle parti erose, sapore mite, odore da nullo in gioventù a forte nei soggetti maturi, più o meno fruttato, non reagisce a contatto con gli alcali.
Reazione macrochimica con la potassa le superfici si tingono di rosso fuoco indipendentemente dalla presenza del pigmento giallo.
Sporata non ottenuta.
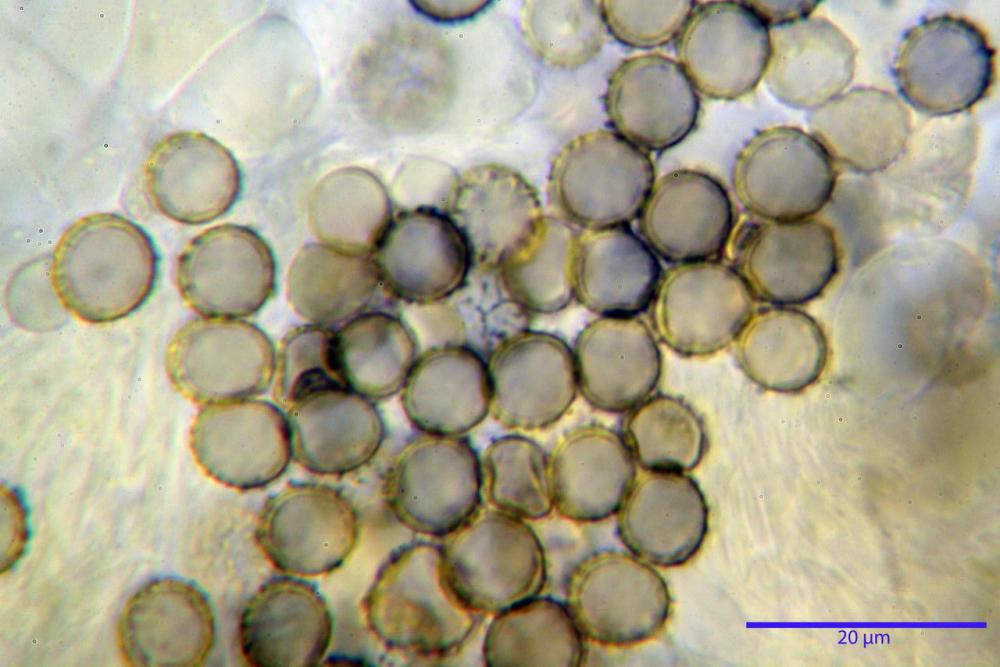
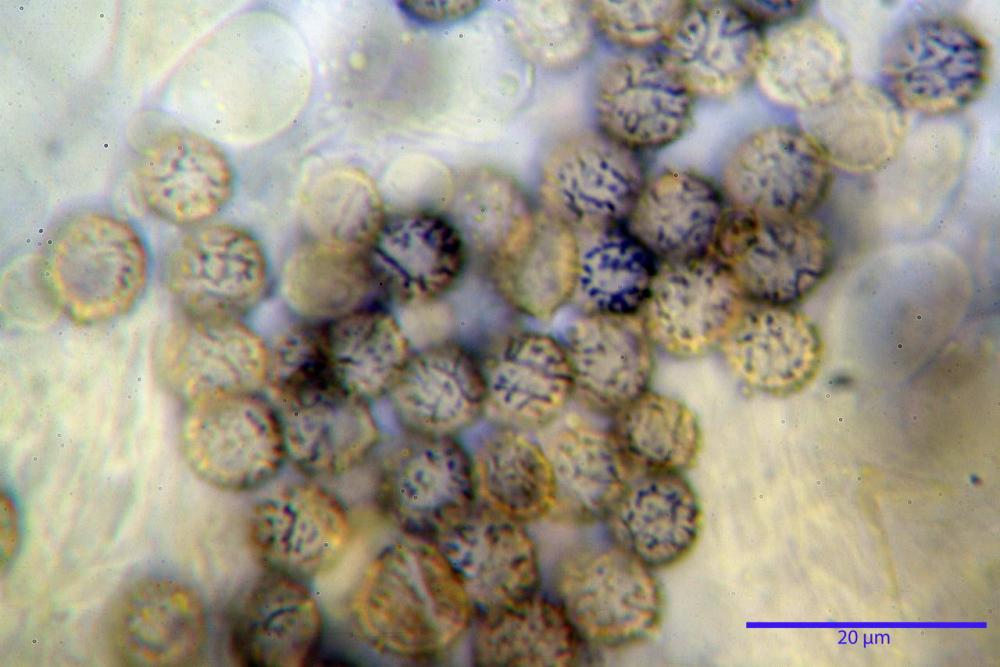
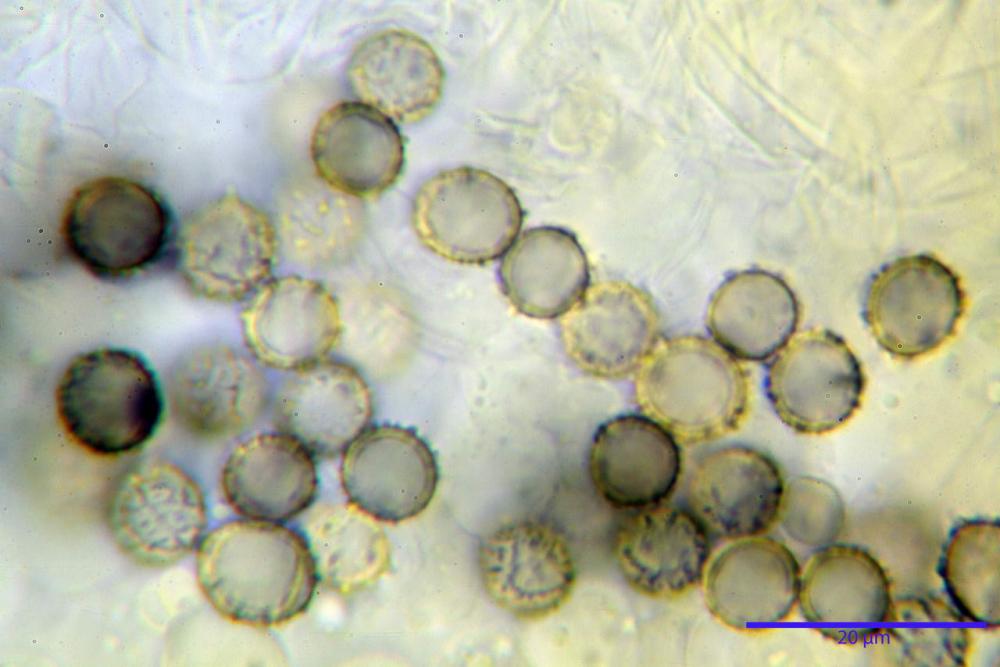
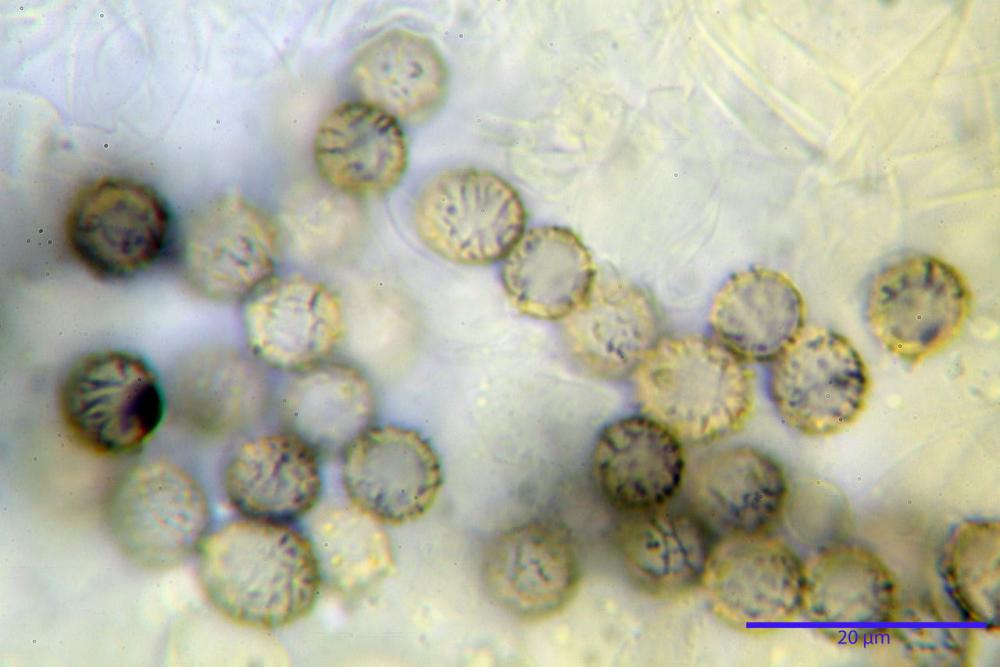
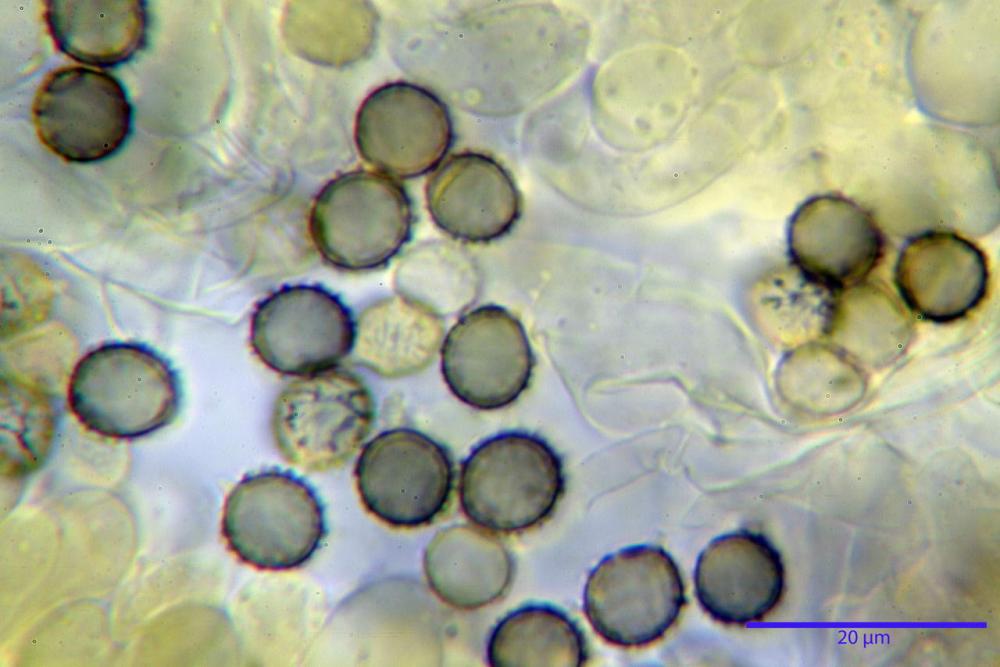
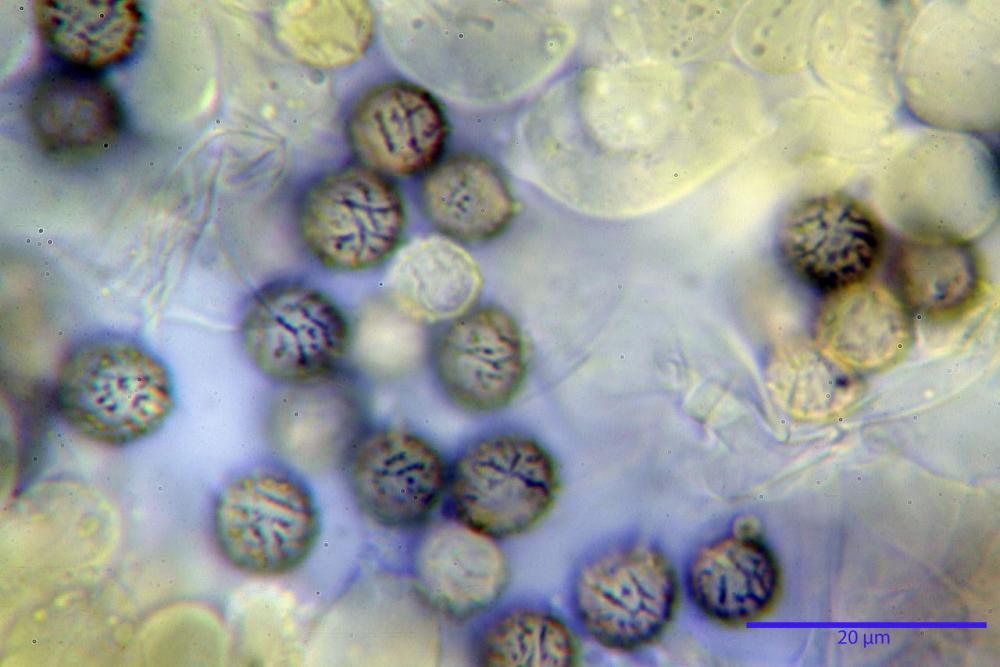
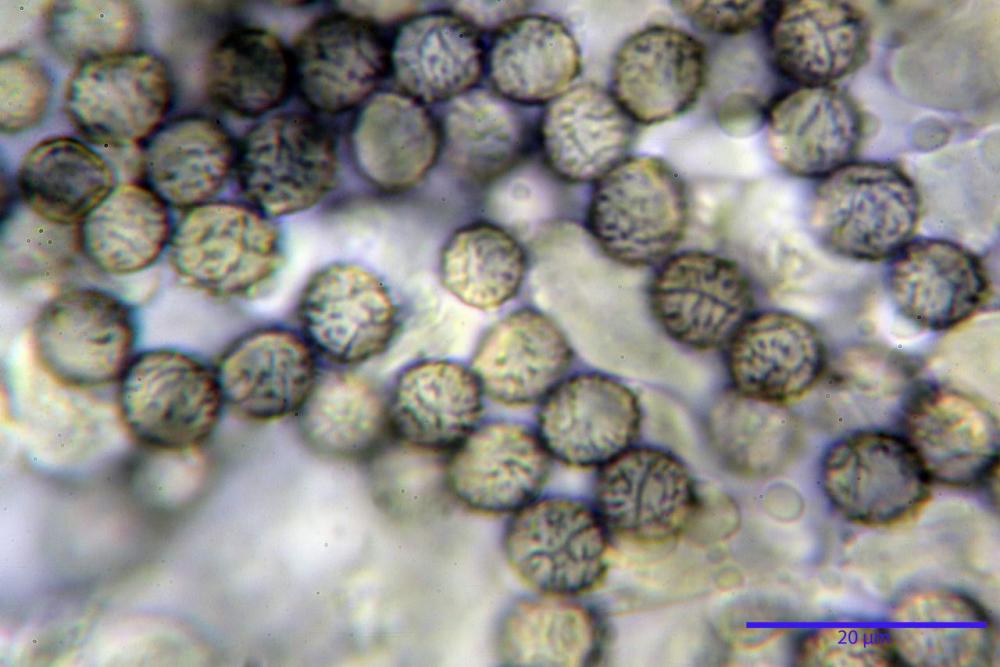
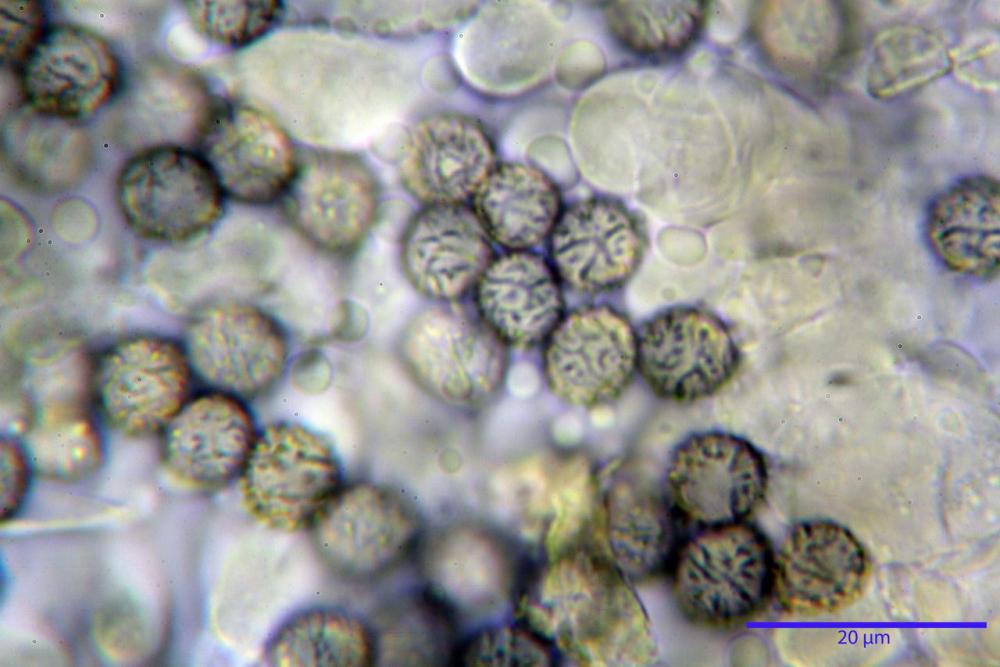
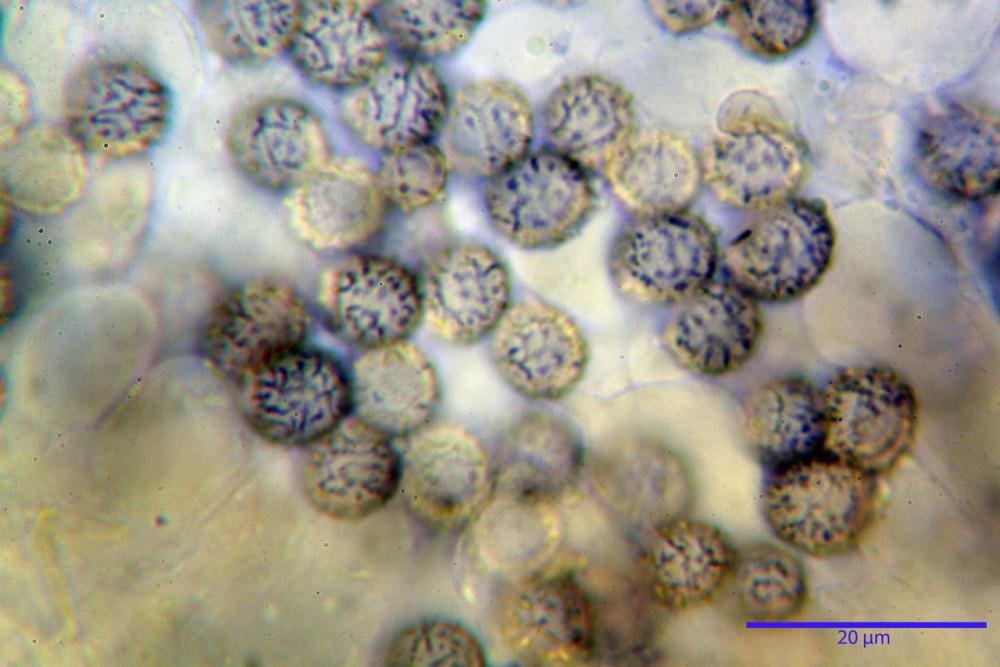
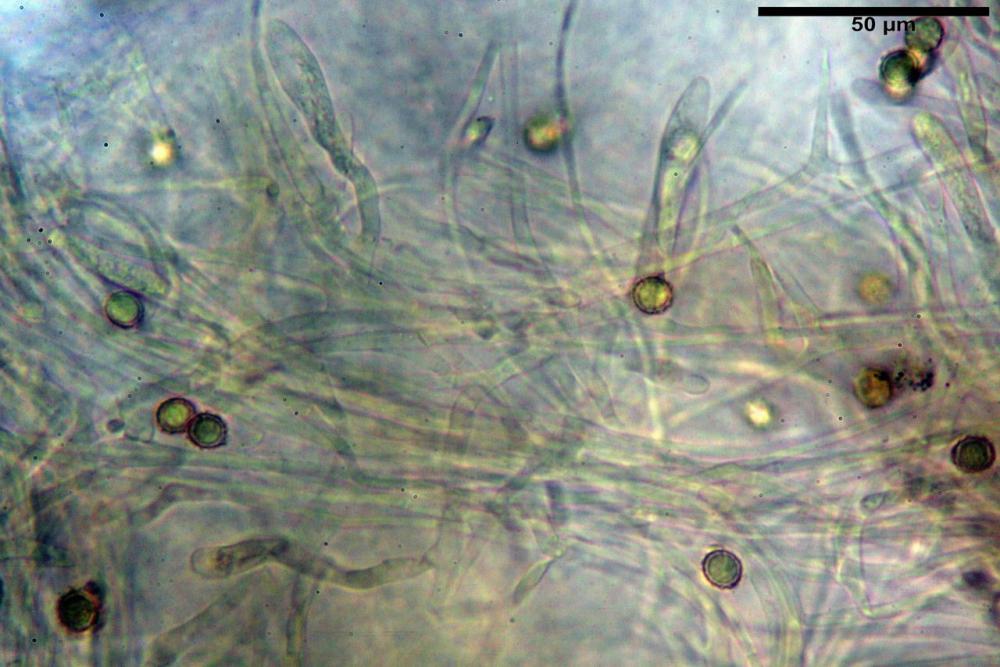
Spore (7,5)8,1-8,6-9,2(10,1) x (6,5)7,2-7,6-8,1(8,8) µm, V. (170,1)220,3 – 266,9-319,1(410,6) µm³, Q. = (1,0)1,0-1,1-1,1(1,2), largamente ovoidali, fino a globose, verrucose-echinulate, crestato-catenulate, subreticolate, con verruche alte fino a 1 µm, con tacca soprailare poco amiloide, con appendice sterigmale in posizione eterotropica.
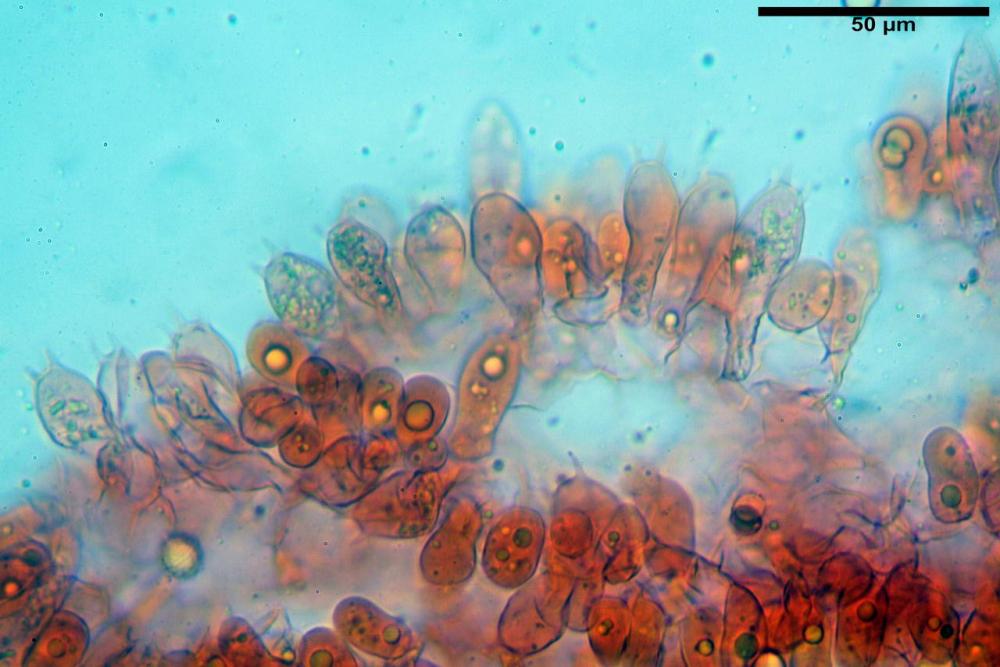
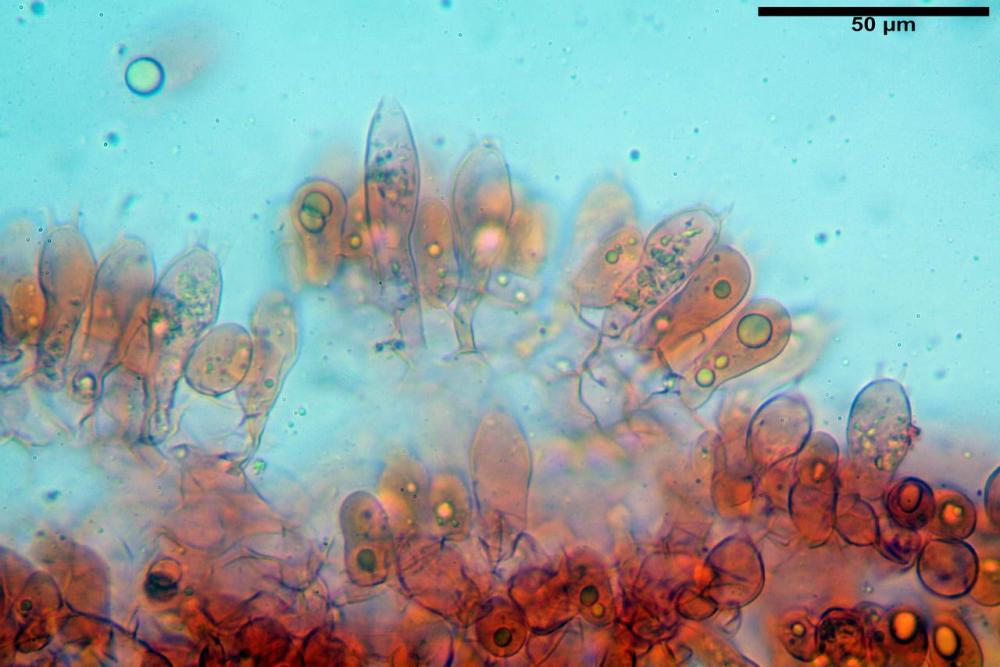
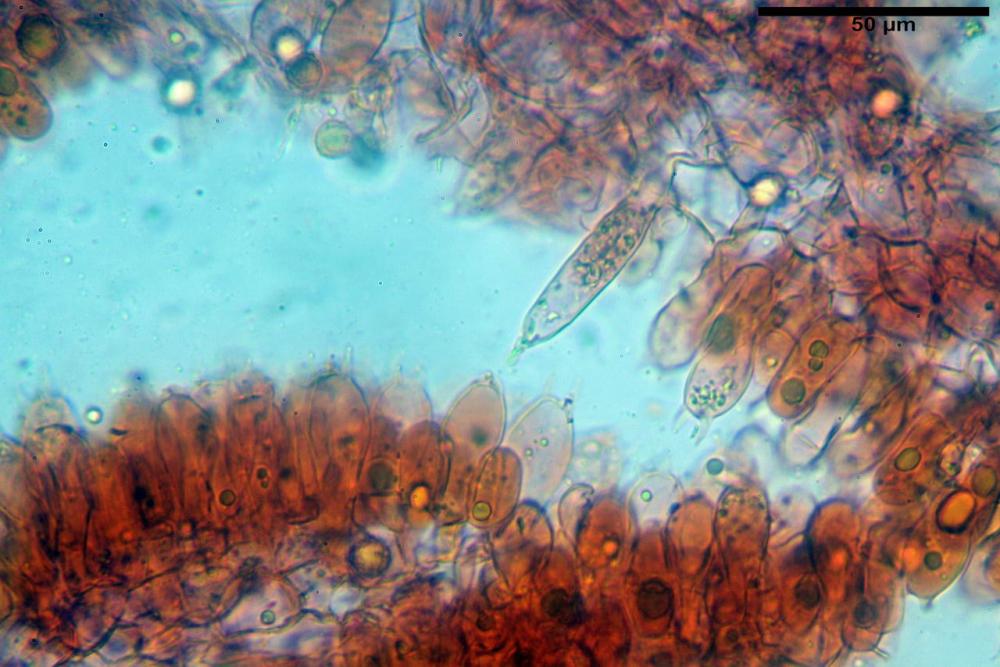
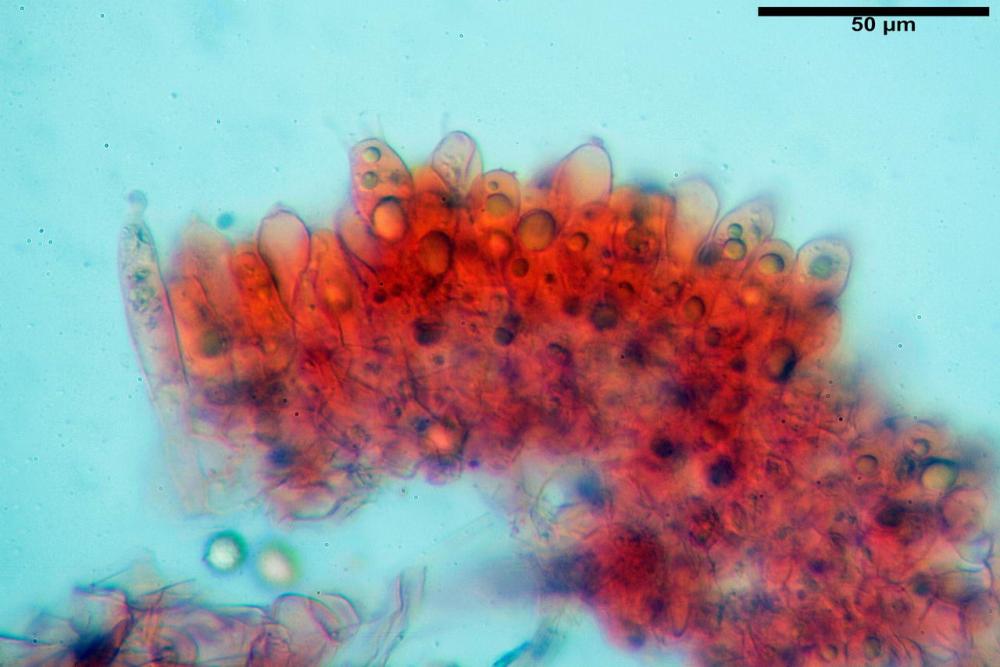
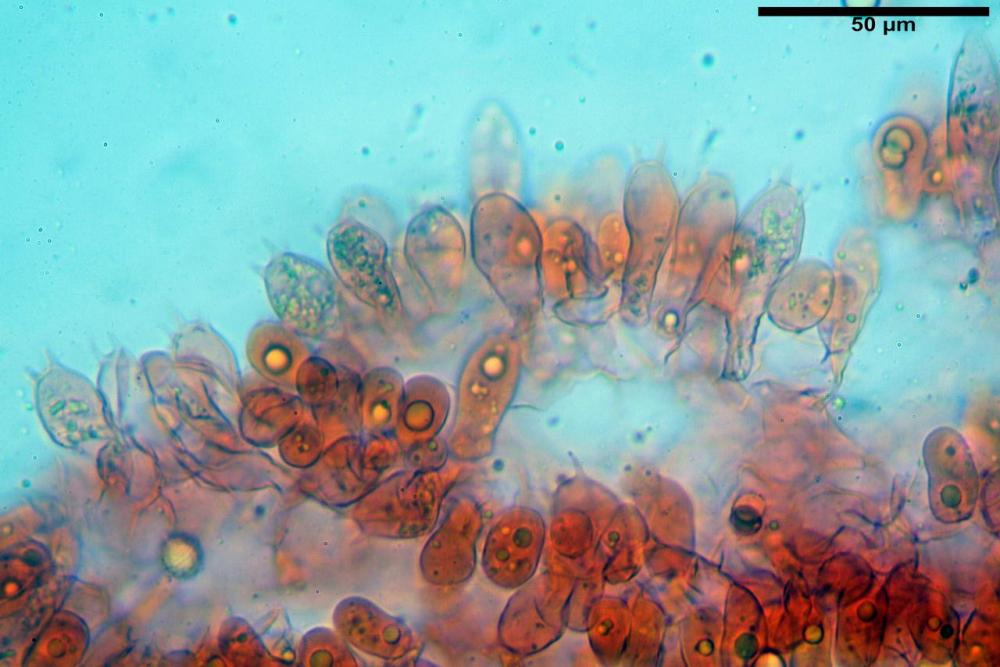
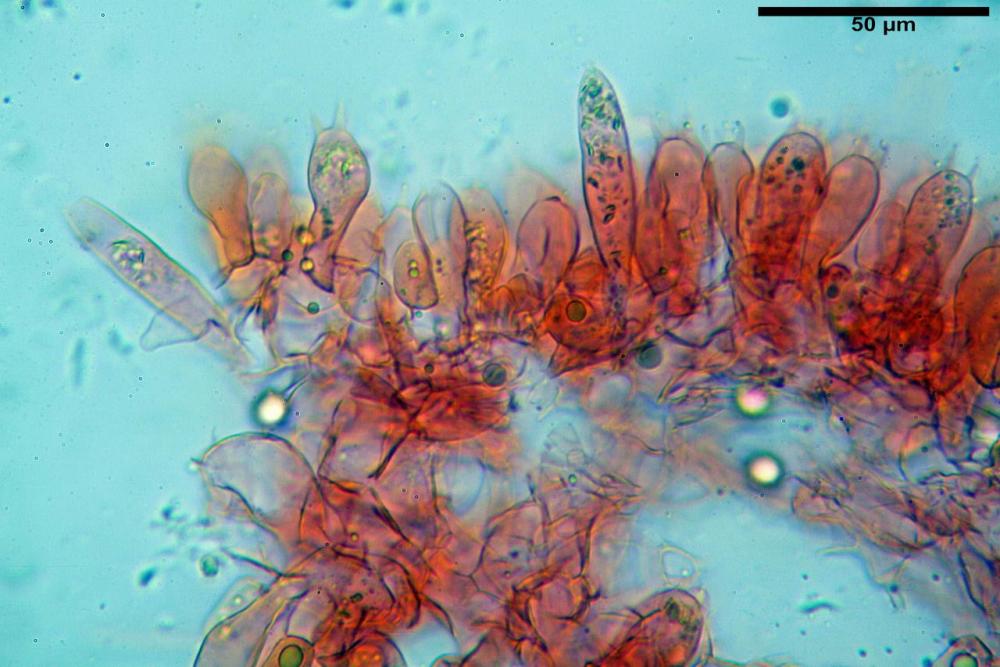
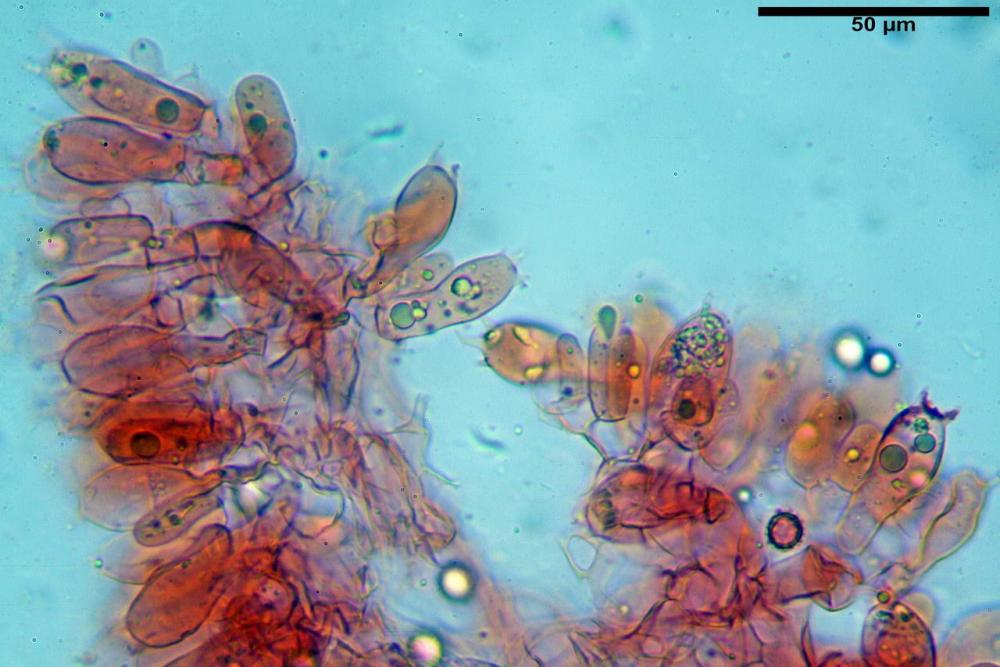
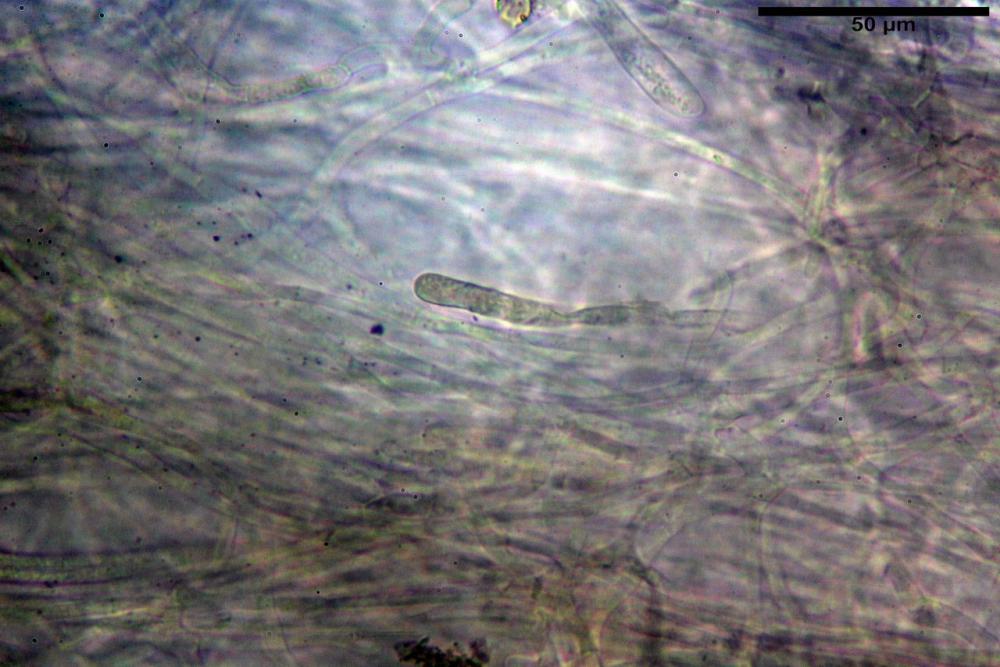
Cistidi imeniali 48,8-58,4-70,3 x 9,3-10,6-11,8 µm, poco numerosi, dispersi tra gli elementi imeniali, da clavati a fusiformi, con sommità a ogiva, alcuni dotati di appendice, con pigmento intracellulare rifrangente di colore giallo.
Basidi (33,9)34,0-38,8-43,6(43,7) x (11,3)11,6-12,7-13,8(14,3) µm, clavati, tetrasporici, con grossi e lunghi sterigmi, alcuni con pigmento intracellulare giallo.
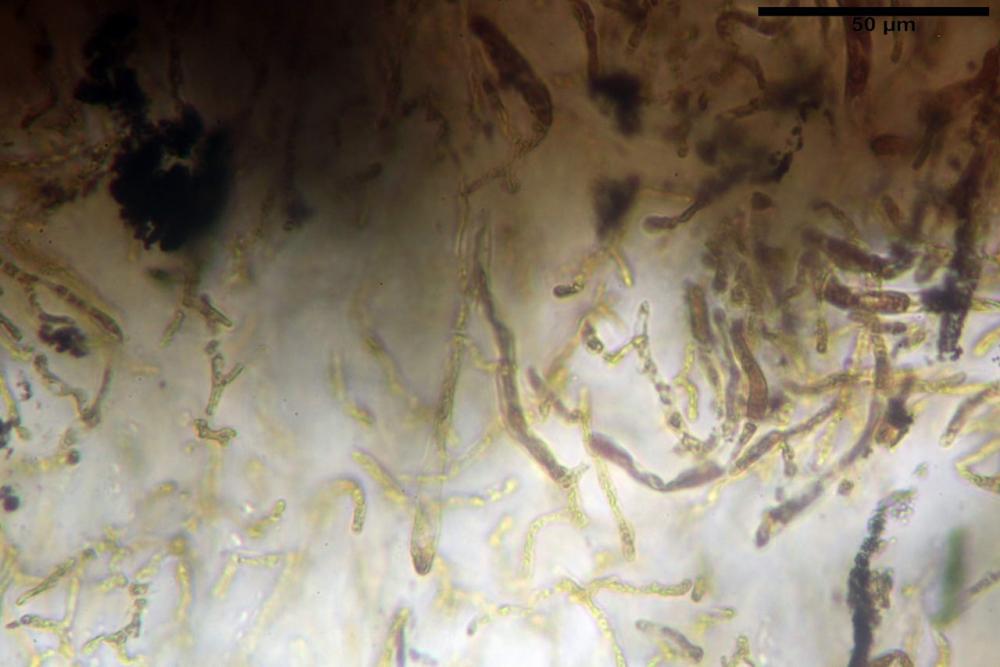
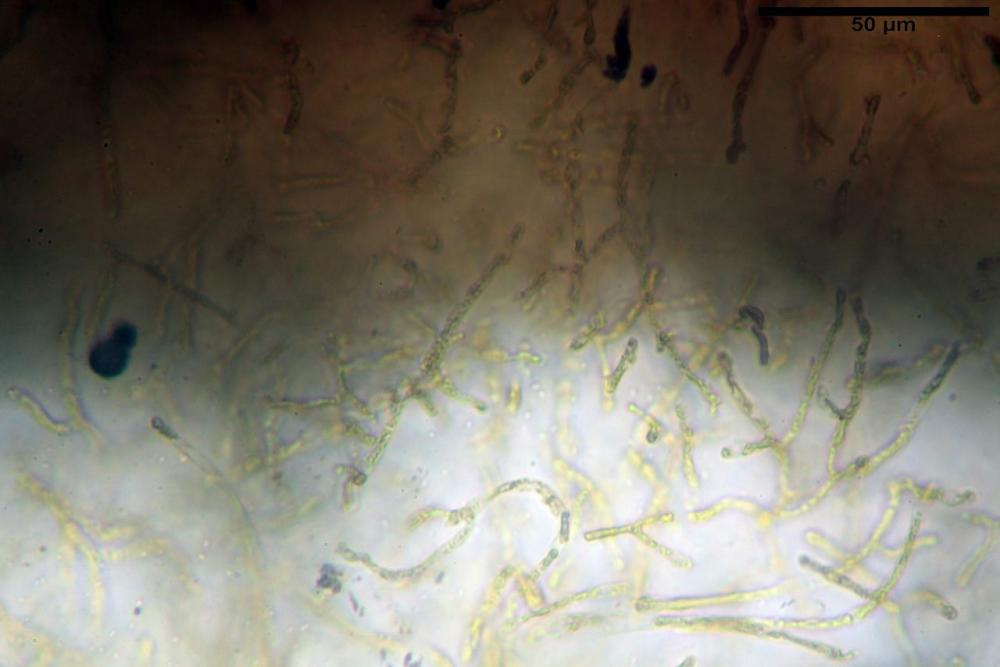
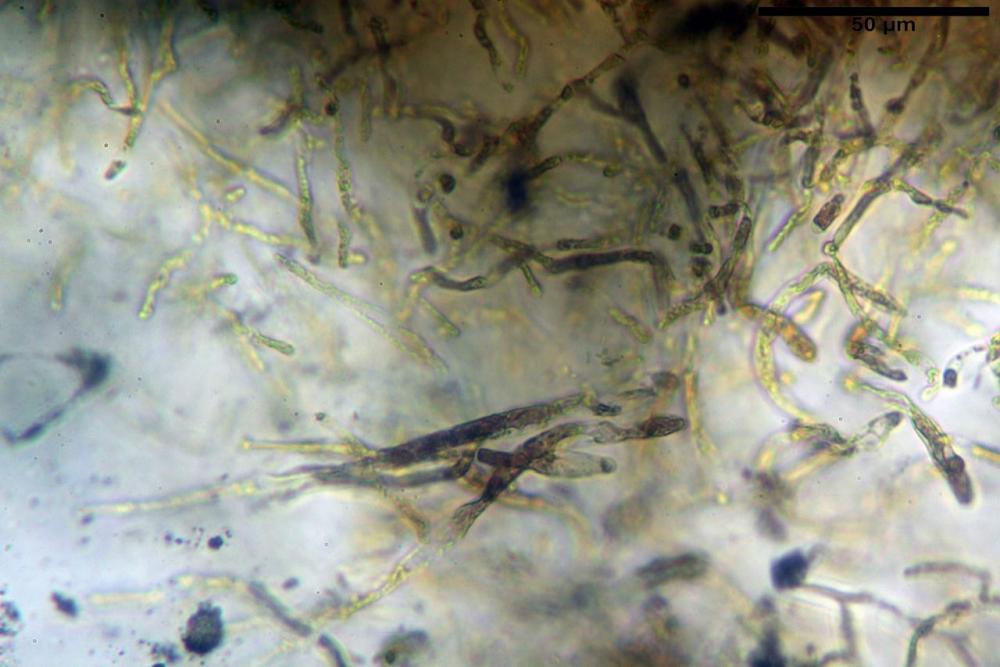
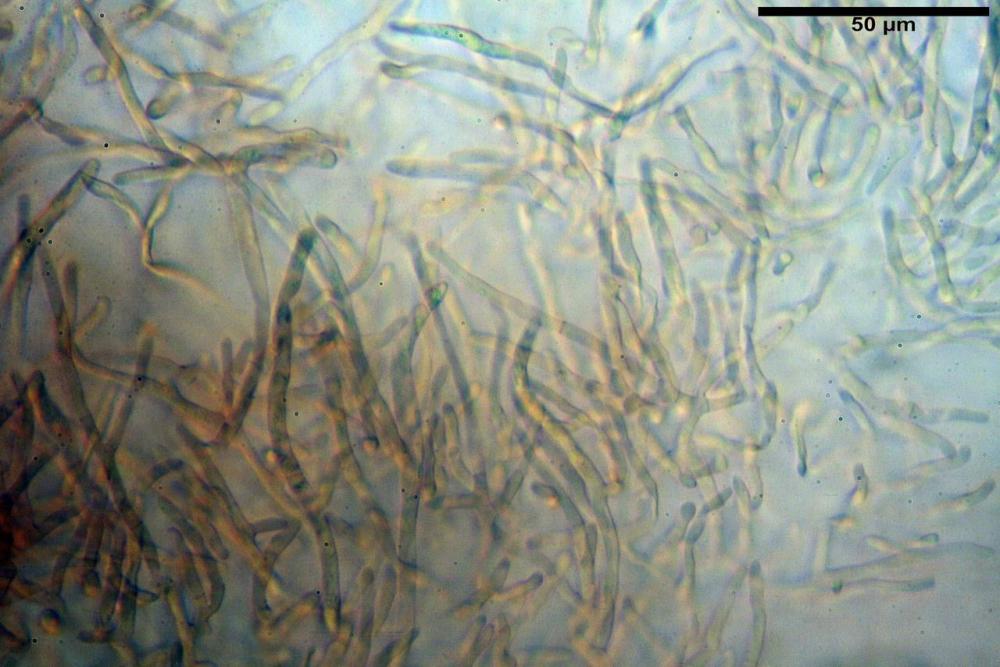
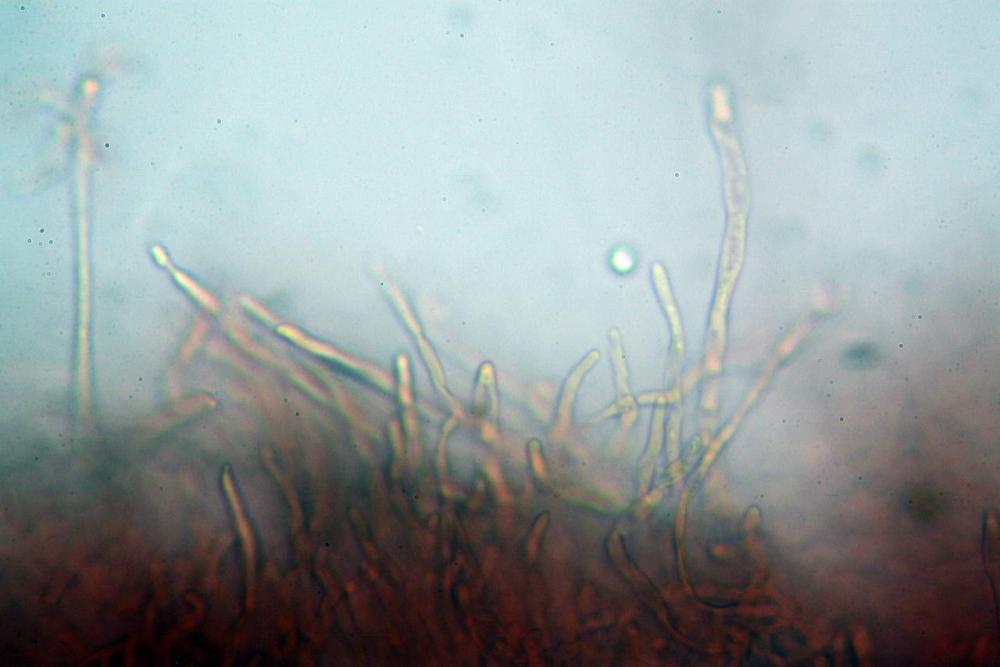
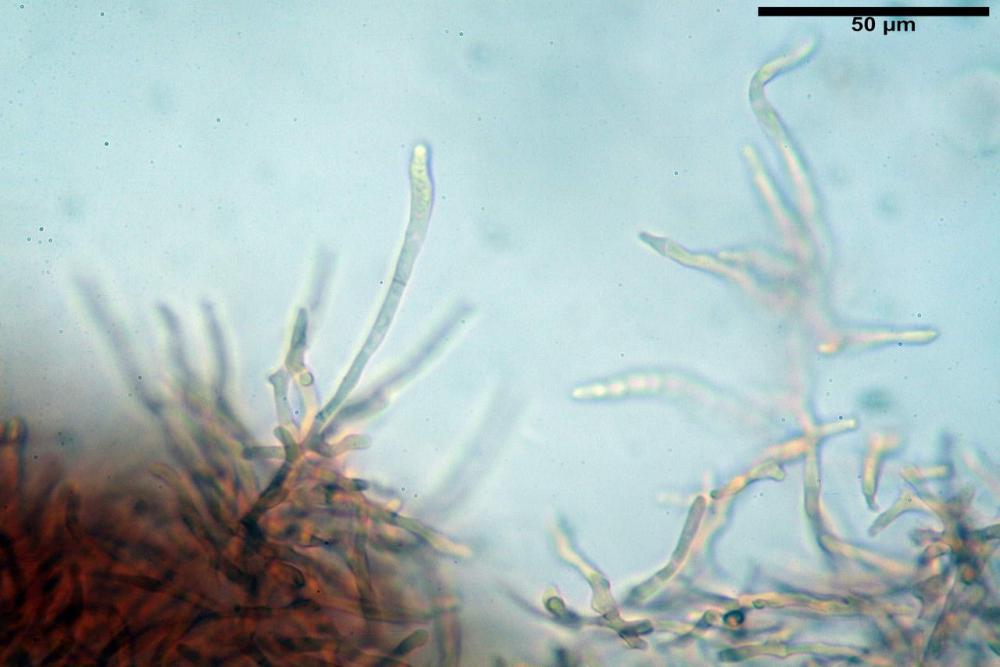
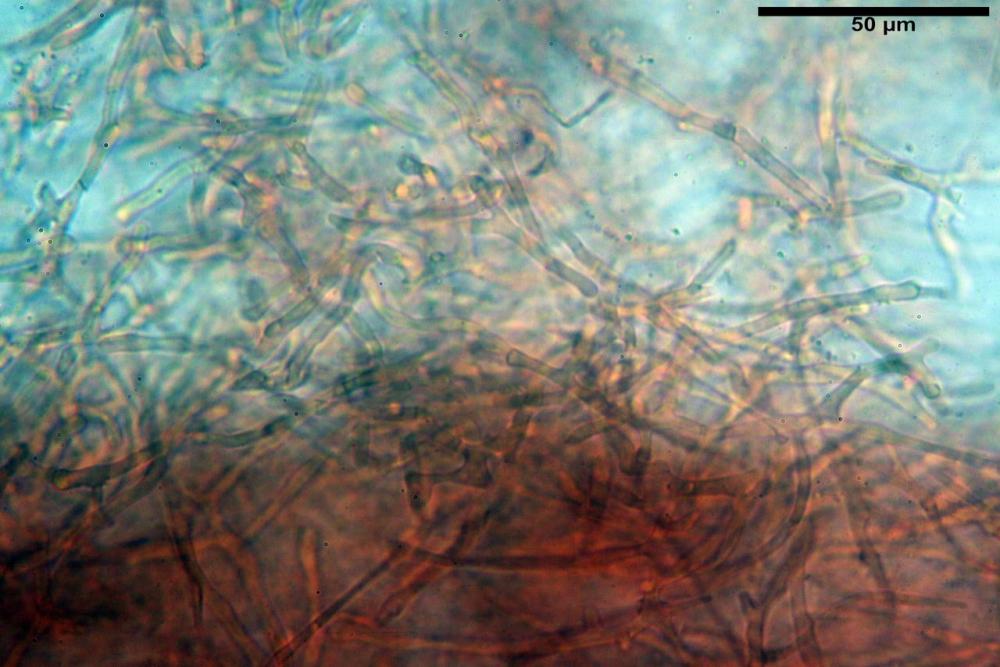
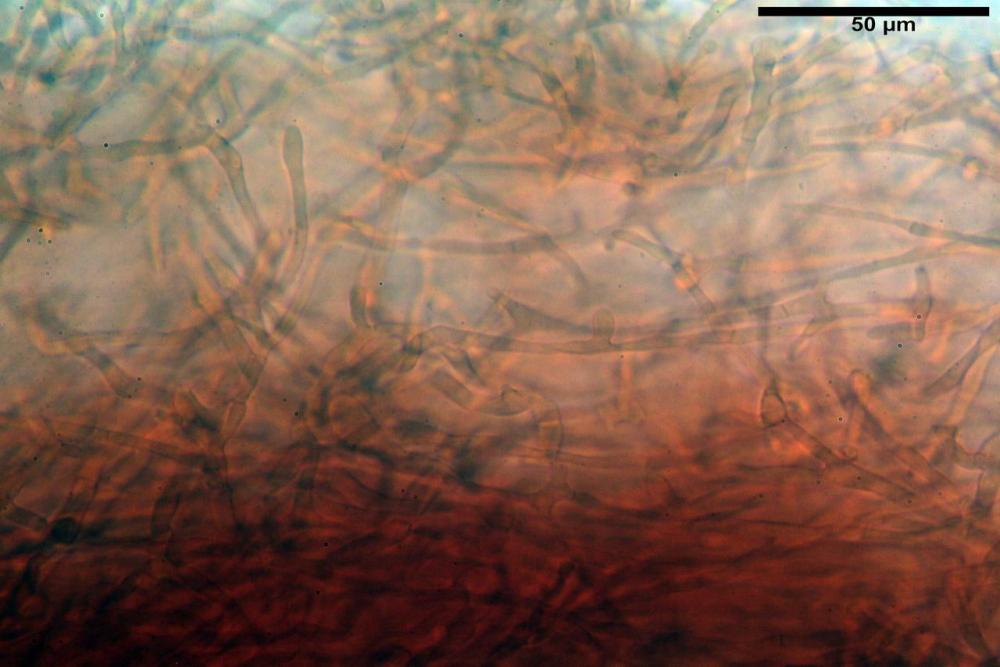
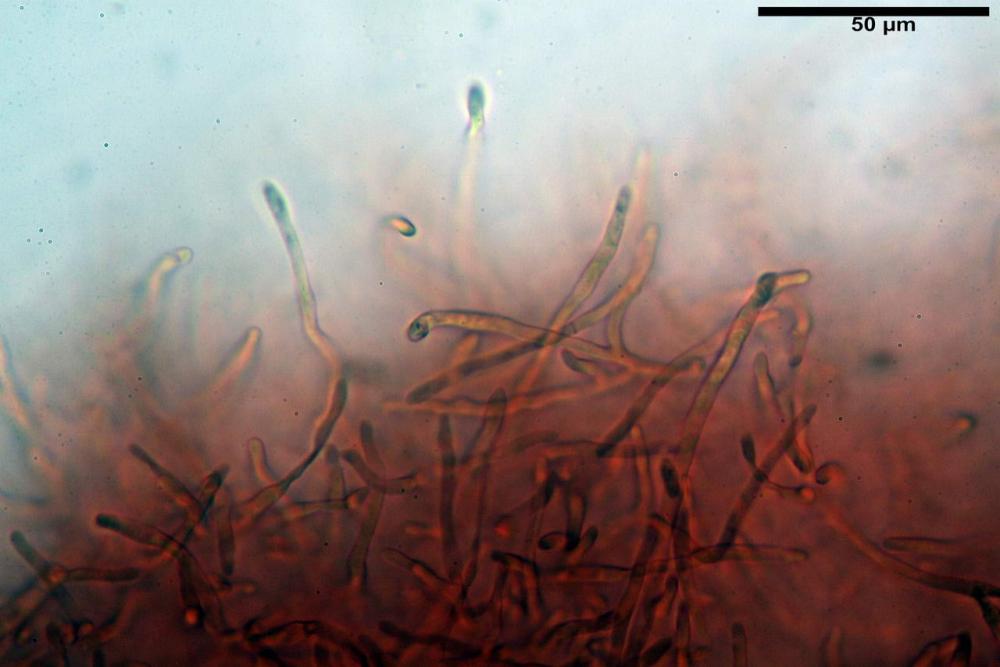
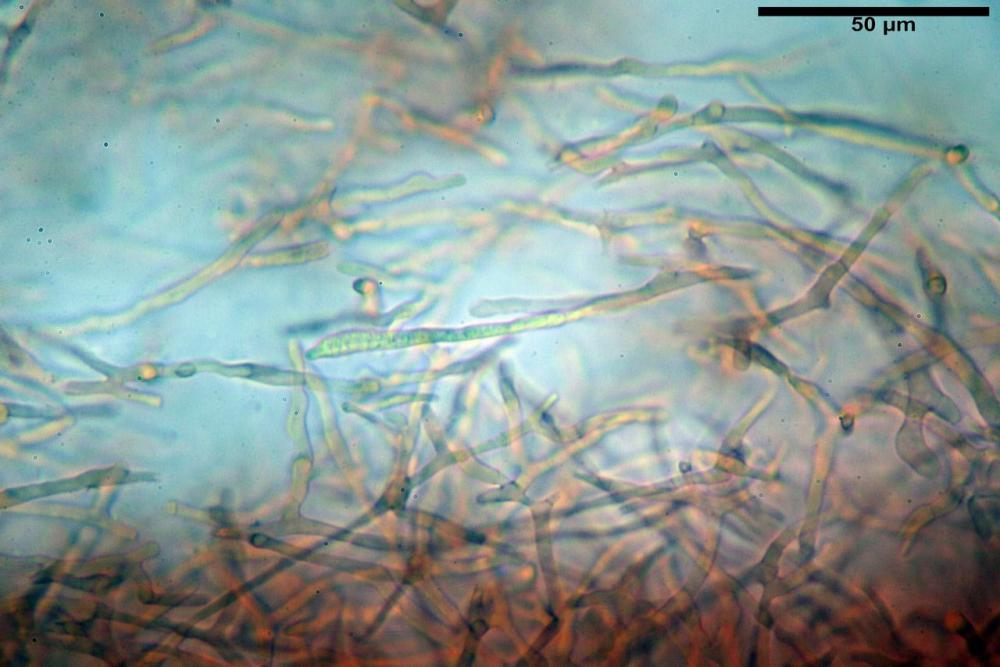
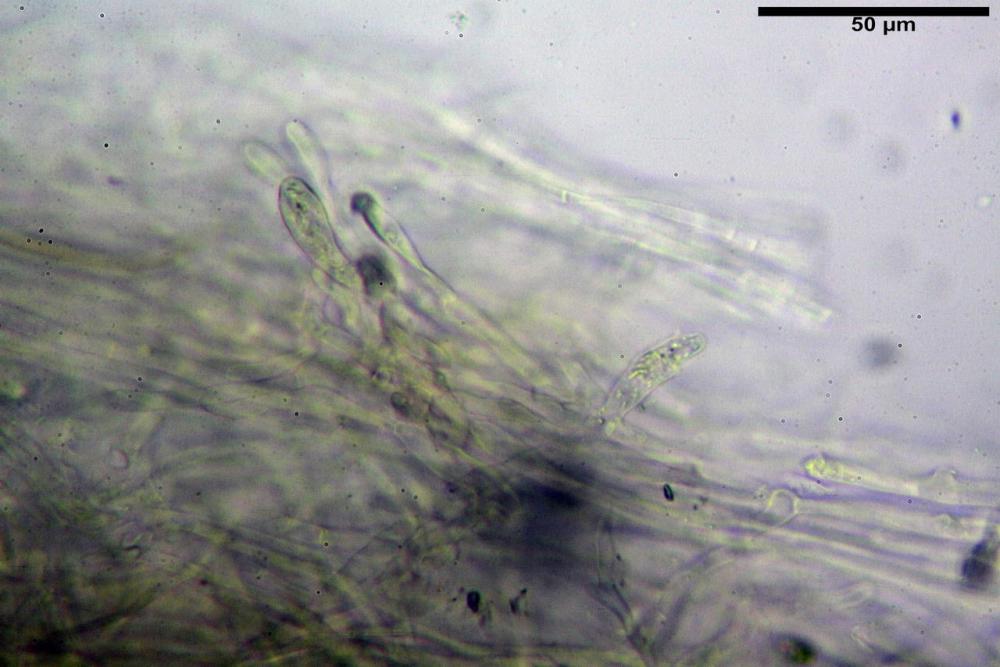
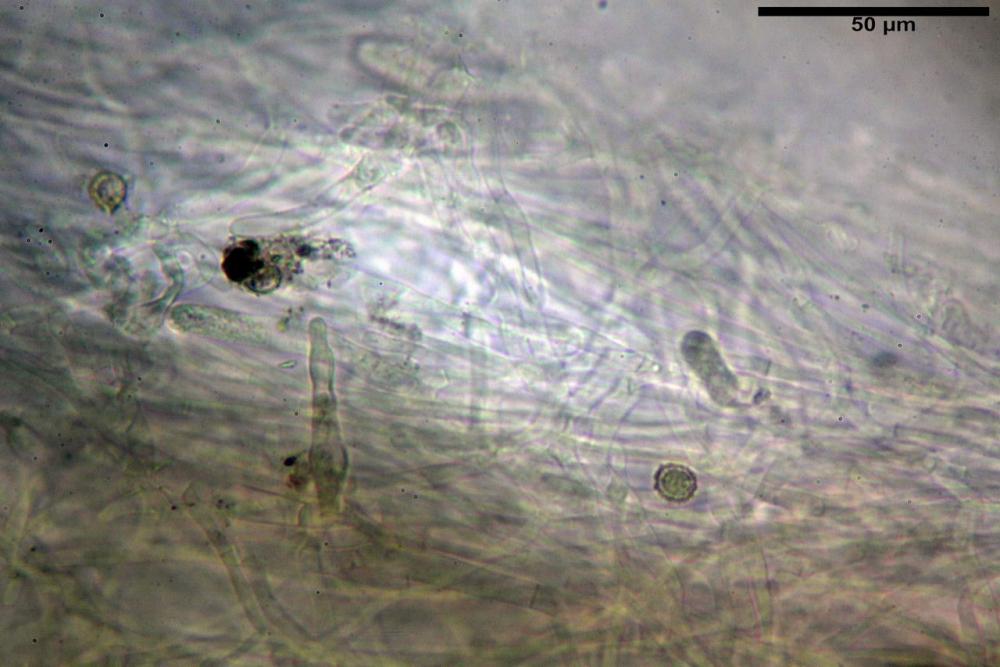
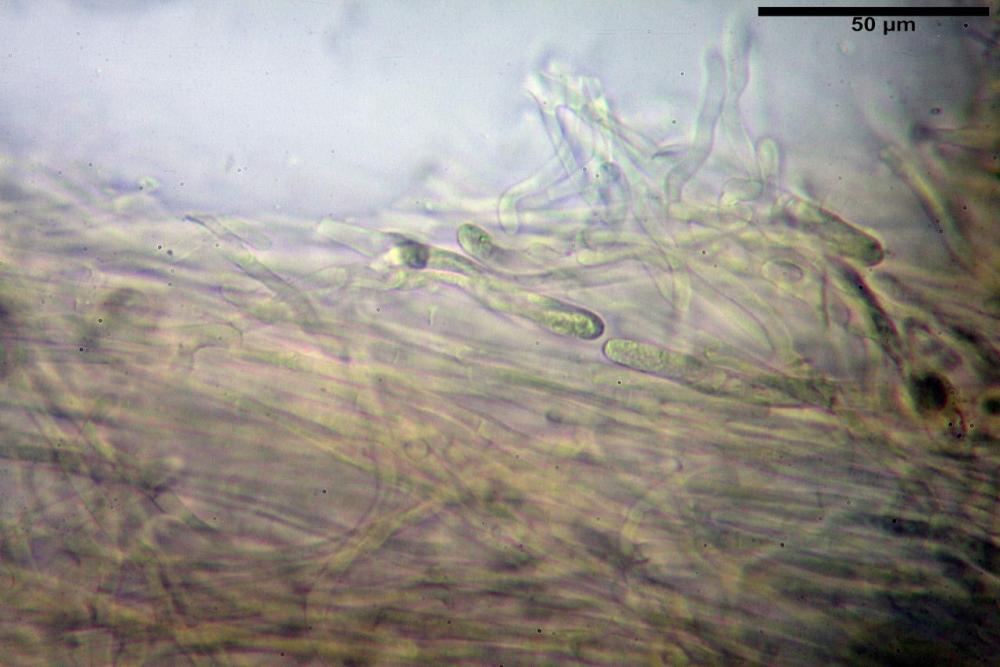
Pileipellis formata da peli cilindracei, ottusi, larghi (2,5)2,6-3,1-3,9(4,1) µm, disposti in un tricoderma o subtricoderma, inframezzate da due elementi cuticolari inguainati da una sostanza amorfa giallo oro; il primo avente una forma cilindrica, con sommità ottusa, di dimensioni 36,0- 60,8-83,7 x 4,25-4,8-6,04 µm, evocante per morfologia dei dermatocistidi, il secondo, piuttosto disperso, con l’estremità attenuata-affilata, avente lo stesso calibro del primo elemento ed evocante per morfologia delle ife primordiali.
Caulopellis composta da ife sinuose, larghe (2,2)2,3 - 3,1-3,8(3,8) µm, ad andamento subparallelo, ialine, frammiste a ife di calibro maggiore, larghe (6,1)6,1 - 7,6-9,4(9,7) µm, lunghe (51,2)51,6 - 60,3-67,6(68,7) µm, più o meno tortuose, con apice clavato, contenenti un abbondante pigmento giallastro rifrangente, evocanti per morfologia dei caulocistidi.
Materiale studiato ed ambiente di raccolta: ITALIA. N. scheda: 7720. Data di ritrovamento: 08/11/2018. Località: Bosco LiLei. Comune: Lizzanello (LE). Coordinate geografiche: 40°18'55.4"N 18°15'16.6"E. Altezza 40 m s.l.m. Habitat: Bosco di Quercus ilex. Raccolta 2018 (n° 7712). Legit. Bruno de Ruvo; Det. Marino Zugna, Bruno de Ruvo, Paolo Picciola, Silva Refellato & Paolo De Martin.
Data inserimento in erbario: 27. 02. 2019
Osservazioni
Abbiamo il piacere di presentare una raccolta composta da esemplari molto belli e tipici di Russula messapica ritrovata nella sua terra d’origine.
Si tratta di una specie molto interessante dalle caratteristiche uniche nell’ambito delle russule europee.
Come riportato da Sarnari (1990) a prima vista richiama una Tenellae per il portamento e la taglia assai ridotta, tuttavia, il suo quadro microscopico rivela l’assenza di dermatocistidi plurisettati, elementi tipici delle Tenellae, che sono rimpiazzati da ciuffi di ife addensate, immerse in un abbondante pigmento amorfo, extramembranario, di colore giallo oro e di origine velare.
Tali elementi cuticolari si presentano con due tipologie: la prima evoca delle ife primordiali per la morfologia lungamente attenuata e l’estremità affilata, la seconda, quella più abbondante, esprime una morfologia pileocistidioide assimilabile ai dermatocistidi dai quali si differenzia per una fine emulsione brunastra con l’idrossido d’ammonio che risulta inerte ai reattivi solfoaldeidici (Sarnari, 1990, 2005).
A contatto con l’idrossido di potassio l’abbondante pigmento giallo produce sulle superfici dei basidiomi una spettacolare e immediata reazione rosso sangue che si attenua fino a scomparire del tutto in una decina di secondi (Sarnari, 1990, 2005).
Inizialmente il suo autore considerò questa specie una sorta di endemismo circoscritto al Salento, ma questa ipotesi fu rivista a seguito di alcune raccolte avvenute al di fuori dell’areale originario: nella località sarda di Cala Gonone in provincia di Nuoro (Sarnari, 2005) e in Spagna presso Gerona (Llistosella & Vidal, 1995; Sarnari, 2005).
Sulla base di queste raccolte Sarnari (2005) riconsiderò la sua russula una entità piuttosto rara, conosciuta per pochissime località della fascia mediterranea costiera (Sardegna e Spagna) e ben distribuita nella penisola salentina nei boschetti sabbiosi di leccio e/o quercia spinosa.
Successivamente, R. messapica è stata ritrovata in altre località italiane: nel bosco della Cernizza a Duino (TS) in Friuli Venezia Giulia (Billa, 2007) e nel Parco Aymerich, Laconi (OR) in Sardegna (Lezzi, 2013).
In Europa è stata recentemente ritrovata in Spagna, nelle isole Baleari (Siquier et al., 2015) e in Grecia nella regione dell’Attica e a Creta (Vidal et al., 2019).
Questi ritrovamenti confermano il suo ampio areale lungo la costa mediterranea, dalla Grecia alla Spagna, dal livello del mare fino ai 600 m di altitudine.
L’arrossamento delle superfici a contatto con gli alcali di R. messapica è una carattere riscontrabile soltanto all’interno di un piccolo gruppo di specie dotate di residui velari.
Tra queste menzioniamo Russula insignis Quél., taxon appartenente alla Sezione Subvelatae del Sottogenere Ingratula, Russula viscida Kudrna e Russula ochroleuca Pers., quest’ultime appartenenti, secondo l’impostazione sistematica di Sarnari (1998, 2005), alla Sezione Viscidinae del Sottogenere Russula. Il confronto tra queste specie con R. messapica si limita soltanto a questa reazione poiché esse sono molto diverse da R. messapica sia sul piano morfologico che su quello sistematico, nonché per una diversa composizione istomorfologica del velo.
Riteniamo, diversamente da quanto sostenuto da Sarnari che R. messapica sia una specie priva di residui velari veri e propri. Secondo la nostra opinione, l’arrossamento agli alcali è da attribuire unicamente alla presenza dell’abbondante pigmento giallo oro che avvolge alcuni elementi della caulopellis e della pileipellis.
Recentissimi studi condotti da Vidal et al. (2019), basati su evidenze molecolari e morfologiche, hanno accertato la conspecificità di Macowanites messapicoides, un taxon angiocarpico descritto dalla Spagna nei pressi di Gerona (Llistosella & Vidal, 1995) con Russula messapica.
Già all’atto della sua descrizione originaria, gli autori Llistosella & Vidal (1995) avevano riconosciuto delle stupefacenti analogie tra la loro specie angiocarpica con quella gimnocarpica di Sarnari, prospettando per entrambe un chiaro nesso di unione filogenetica fra le russule gasteroidi e quelle agaricoidi.
Qualche anno dopo, Martin et al. (1999), effettuando delle analisi molecolari delle due specie nelle regioni ITS e l’IGS, avevano concluso che esse sono monomorfiche per tutti i markers tranne che per l’IGS e che questa limitata differenziazione genetica stava a indicare una stretta relazione tra le due specie.
Pur mantenendo distinte le due entità, gli autori di questo studio suggerivano la possibilità che Macowanites messapicoides fosse una forma gasteroide derivata da Russula messapica.
Più tardi, anche Miller et al. (2006) considerarono questa specie una forma angiocarpica conspecifica di Russula messapica, tuttavia non formalizzarono questa sinonimia.
Solo recentemente, nell’ambito di un’ampia revisione nomenclaturale delle Russulaceae gasteroidi (sequestrate Russulaceae), Elliott & Trappe (2018) hanno provveduto a trasferire Macowanites messapicoides nel Genere Russula.
Come sopra riferito, il lungo iter nomenclaturale e sistematico si è concluso con lo studio di Vidal et al. (2019) basato su sequenze di quattro markes del DNA ottenute da diverse raccolte di M. messapicoides e R. messapica provenienti da diversi paesi dell’area mediterranea.
I risultati non hanno evidenziato delle differenze significative tra i due taxa, pertanto, la forma angiocarpica è stata differenziata a livello varietale dalla forma gimnocarpica per il diverso habitus.
Morfologicamente, R. messapica var. messapicoides è da considerare a tutti gli effetti una forma angiocarpica in miniatura di R. messapica var. messapica, dove le lamelle e lo stipite sono progressivamente trasformati in alveoli e columella (Vidal et al., 2019)
Per quanto concerne gli altri caratteri, così come indicato da Llistosella & Vidal (1995) le due varietà presentano lo stesso colore, la stessa reazione del peridio agli alcali e il medesimo odore. Microscopicamente, entrambe possiedono un abbondante essudato amorfo pigmentato di giallo oro e sono prive di autentici dermatocistidi.
Infine, i risultati dello studio di Vidal et al. (2019) hanno dimostrato che le due varietà di R. messapica sono significativamente associate al subsezione Puellarinae, mostrando una stretta relazione con R. cessans, R. odorata, R. puellaris e R. versicolor.
Questi risultati sono in linea con il pensiero sistematico di Sarnari.
Bibliografia
Billa L.D. – 2007: Alcune specie di russula rare per il Friuli Venezia Giulia. Bollettino del Centro Micologico Friulano. Udine: 28-31.
Lezzi T. – 2013: Segnalazione di un ritrovamento di Russula messapica in Sardegna (Italia) e confronto con le specie simili munite di velo. Bollettino dell’Associazione Micologica ed Ecologica Romana. (89): 44-47.
Llistosella J. & J.M. Vidal – 1995: Due nuove specie di Russulales Gasteroidi della regione mediterranea. Rivista di Micologia. (2): 149-162.
Martín M.P., N. Högberg & J. Llistosella – 1999: Macowanites messapicoides, a hypogeous relative of Russula messapica. Mycological Research (103): 203-208.
Miller S.L., E. Larsson, K.H. Larsson, A. Verbeken & J. Nuytinck – 2006: Perspectives in the new Russulales. Mycologia 98: 960–970.
Montecchi A. & M. Sarasini – 2000: Funghi ipogei d’Europa. A.M.B. Fondazione Centro Studi Micologici.
Siquier J.L., J.C. Salom, J. Espinosa, F. Esteve-Raventós, J. Llistosella & S. Gomes – 2015: Contribució al coneixement micològic de les illes balears (Espanya). XXI. Revista Catalana de Micologia. (36): 59-88.
Sarnari M. – 1989: Russula messapica Sarnari sp. nov. Bollettino dell’Associazione Micologica ed Ecologica Romana. (18): 12-13.
Sarnari M. – 1990: Russula nuove o interessanti dell’Italia centrale e mediterranea. IX contributo. Rivista di Micologia. (1): 42-43.
Sarnari M. – 1990: Russulae nuove o interessanti dell’Italia centrale e mediterranea. X contributo. Rivista di Micologia. (2): 158-169.
Sarnari M. – 1998: Il Genere Russula in Europa. Tomo 1. A.M.B. Fondazione Centro Studi Micologici. Trento.
Sarnari M. – 2005: Il Genere Russula in Europa. Tomo 2. A.M.B. Fondazione Centro Studi Micologici. Trento.
Vidal J.M., P. Alvarado, M. Loizides, G. Konstantinidis, P. Chachula, P. Mleczko, G. Moreno, A. Vizzini, M. Krakhmalnyi, A. Paz, J. Cabero, V. Kaounas, M. Slavova, B. Moreno-Arroyo & J. Llistosella – 2019: A phylogenetic and taxonomic revision Russulaceae in Mediterranean and temperate Europe. Persoonia (42): 127-185.
Photo by: Bruno de Ruvo