Questa discussione è bloccata
Questa discussione è bloccata
Homophron spadiceum (P. Kumm.) Örstadius & E. Larss., in Örstadius, Ryberg & Larsson (2015)
Basionimo: Agaricus spadiceus Schaeff., Fung. Bavar. Palat.1: tab. 60 (1762)
Sinonimi: Agaricus compactus γ sarcocephalus (Fr.) Fr., Syst. mycol. (Lundae) 1: 290 (1821).
Agaricus fuscescens Batsch, Elench. fung. (Halle): 77 (1783).
Agaricus hygrophilus Pers., Mycol. eur. (Erlanga) 3: 113 (1828).
Agaricus sarcocephalus Fr., Observ. mycol. (Havniae) 1: 51 (1815).
Agaricus spadiceus Schaeff., Fung. bavar. palat. nasc. (Ratisbonae) 4: 27 (1774).
Agaricus spadiceus Schaeff., Fung. bavar. palat. nasc. (Ratisbonae) 1: tab. 60 (1762).
Coprinarius sarcocephalus (Fr.) P. Cout. Eubasidiom. Lusit. Herb. Univ. Olisip. 88. 1919.
Drosophila sarcocephala (Fr.) Quél., Enchir. fung. (Paris): 116 (1886).
Drosophila spadicea (P. Kumm.) Quél., Enchir. fung. (Paris): 116 (1886).
Drosophila spadicea var. hygrophila (Pers.) Quél., Enchir. fung. (Paris): 117 (1886).
Pilosace spadiceus (P. Kumm.) Kuntze, Revis. gen. pl. (Leipzig) 3(2): 504 (1898).
Pratella spadicea (P. Kumm.) J. Schröt., in Cohn, Krypt.-Fl. Schlesien (Breslau) 3.1(33–40): 568 (1889).
Psathyra sarcocephala (Fr.) G. Bertrand, Bull. Soc. mycol. Fr.17: 277 (1901).
Psathyra spadicea (P. Kumm.) Singer, Annls mycol.34 (4/5): 339 (1936).
Psathyrella sarcocephala (Fr.) Singer, Lilloa 22: 468 (1951) [1949].
Psathyrella spadicea (P. Kumm.) Singer, Lilloa 22: 468 (1951) [1949].
Psilocybe sarcocephala (Fr.) Gillet, Hyménomycètes (Alençon): 586 (1878).
Psilocybe cookei Sacc., Syll. fung. (Abellini) 5: 1043 (1887).
Psilocybe spadicea P. Kumm., Führ. Pilzk. (Zerbst): 71 (1871).
Sistematica: Psathyrellaceae, Agaricales, Agaricomycetidae, Agaricomycetes, Agaricomycotina, Basidiomycota, Fungi
Descrizione macroscopica
Basidiomi di taglia media, carne scarsa, acquosa, a crescita aggregata, terricoli, igrofani.
Pileo con diametro fino a 60 mm, all’inizio emisferico, quindi convesso, infine sub-applanato con o senza un basso umbone ottuso, margine a lungo involuto. Cuticola più o meno liscia, di colore beige, ocracea nella porzione discale; margine intero, non striato, più o meno pelosetto. Velo non appezzabile.
Lamelle emarginate, sinuose, all'inizio beige-ocracee, via via più scure, infine marrone tabacco, intercalate da 3 (4) lamellule, filo lamellare bianco e finemente crenulato.
Stipite 30-80 x 3-10 mm, cilindrico, diritto o sinuoso, di colore bianco, alle volte con lievi tonalità beige, nella porzione apicale rivestito finemente da pruina e fibrille concolori, base ricoperta da feltro miceliare bianco; all’inizio pieno, poi fistoloso, base di uguale spessore o soltanto leggermente allargata.
Carne poco consistente, acquosetta nel pileo, fibrosetta nello stipite, di colore bianco ghiaccio in ogni sua parte, leggermente ingrigente nella porzione pileica. In exsiccata, i basidiomi, assumono una colorazione molto scura (marrone tabacco).
Odore e sapore non significativi.
Descrizione microscopica
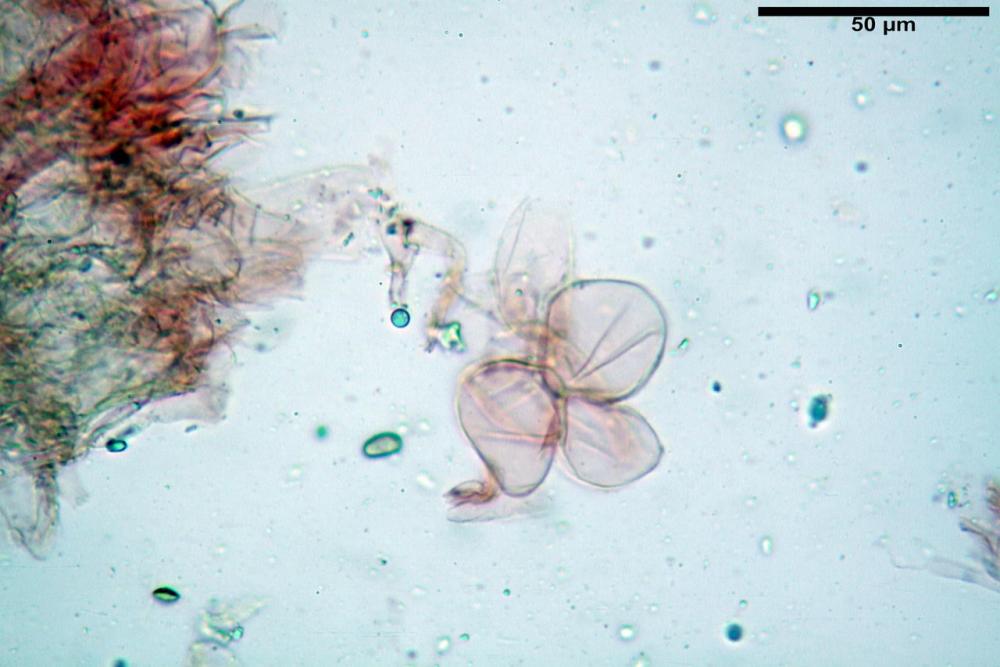
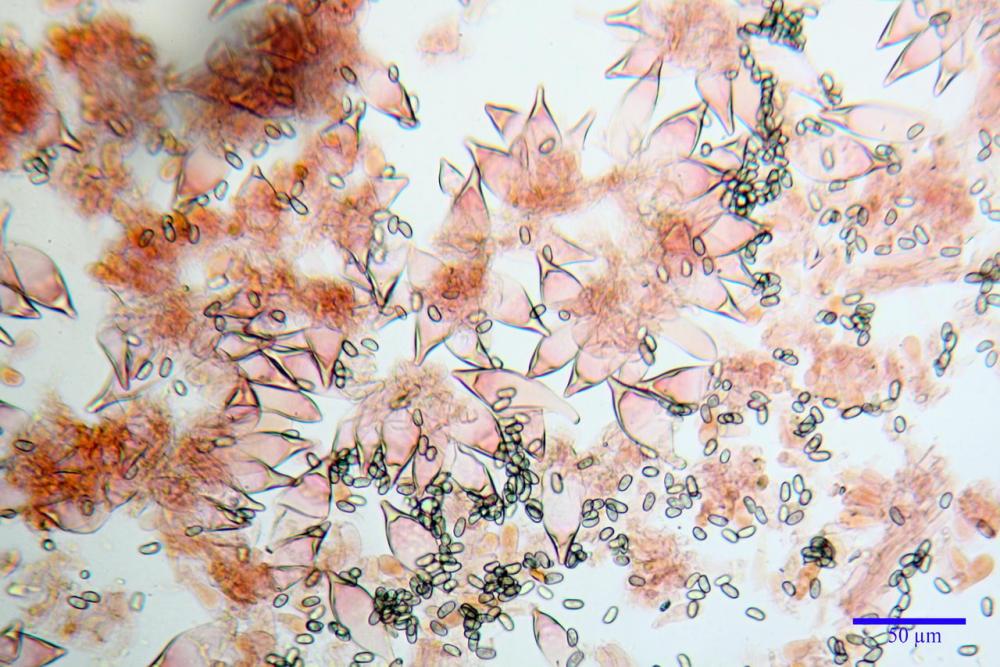
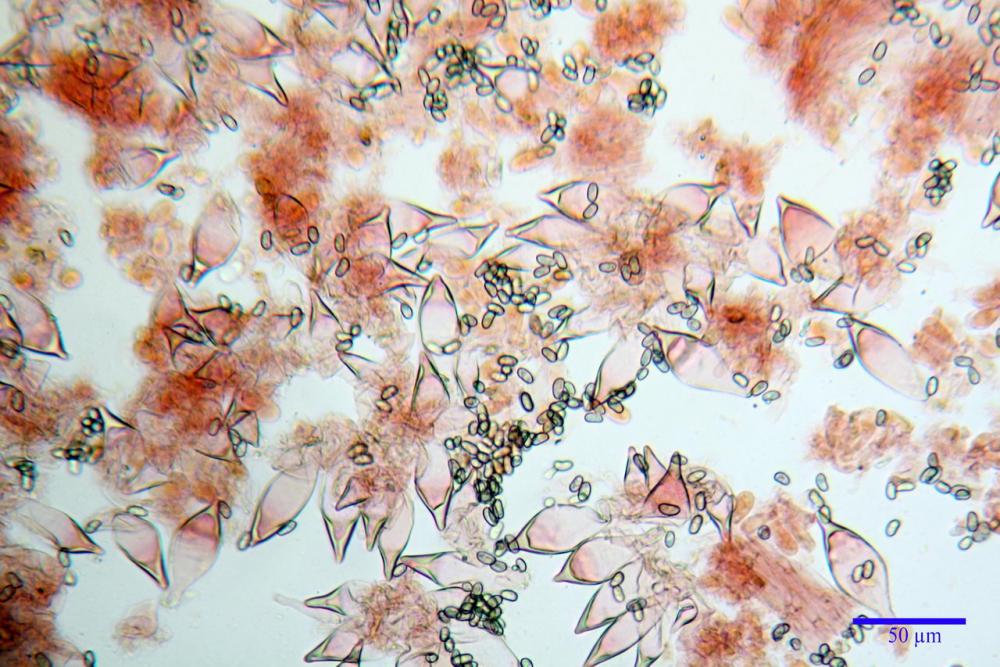
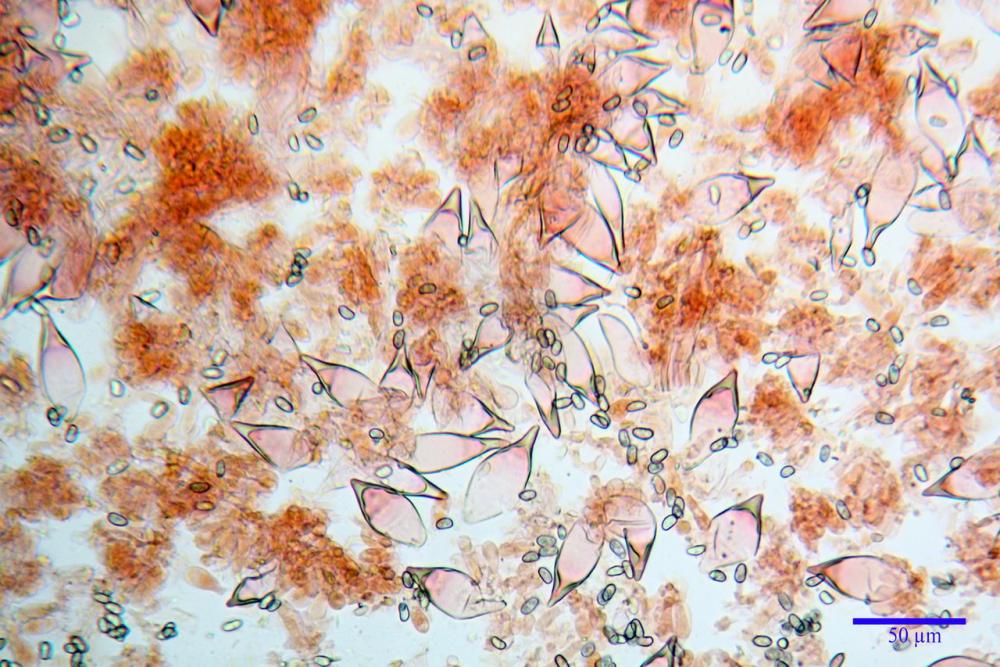
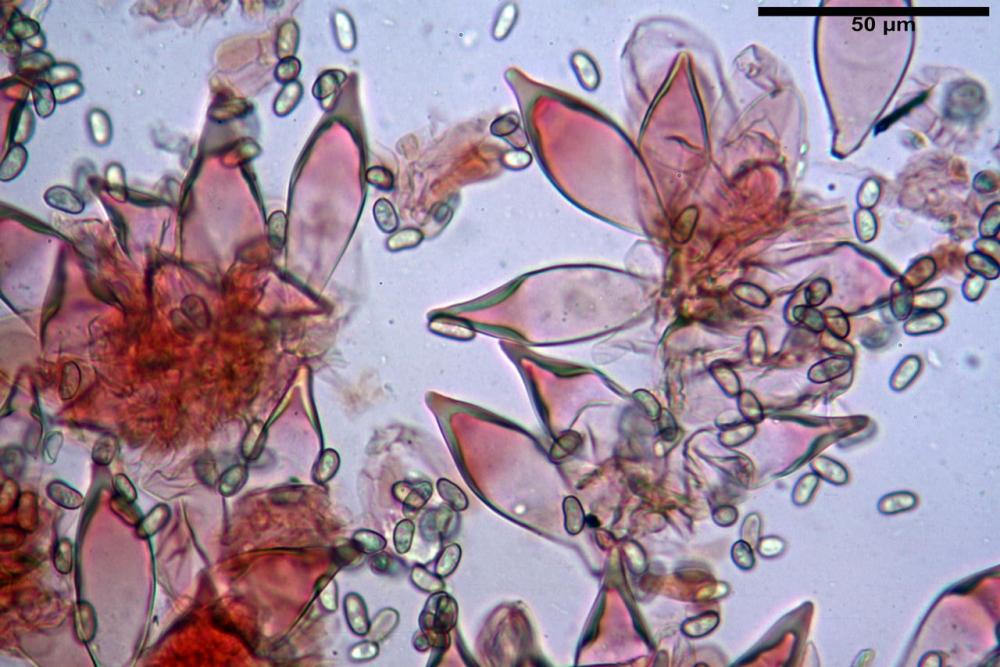
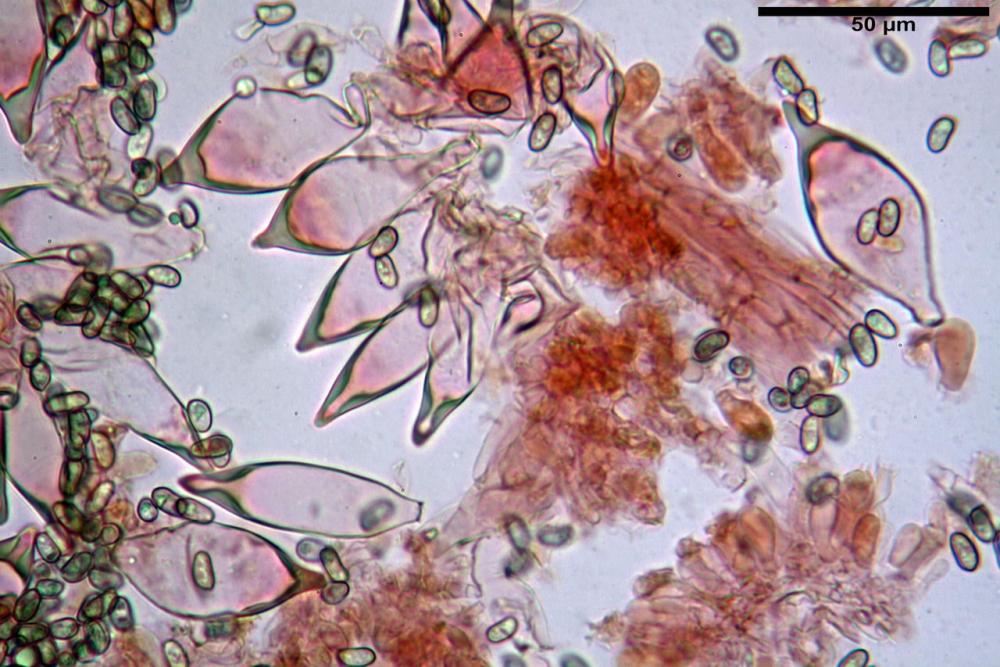
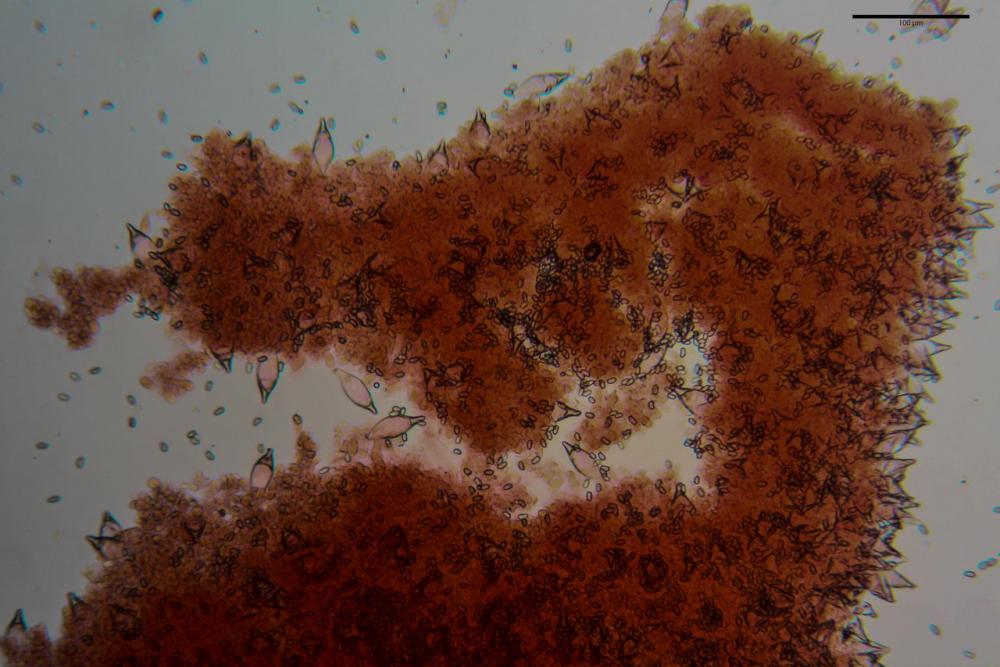
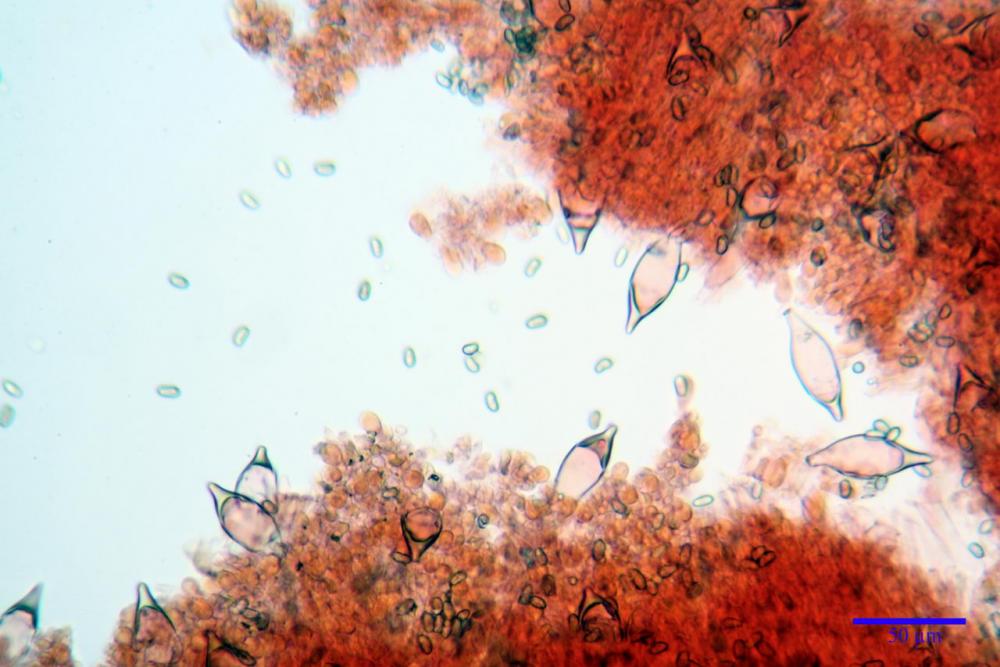
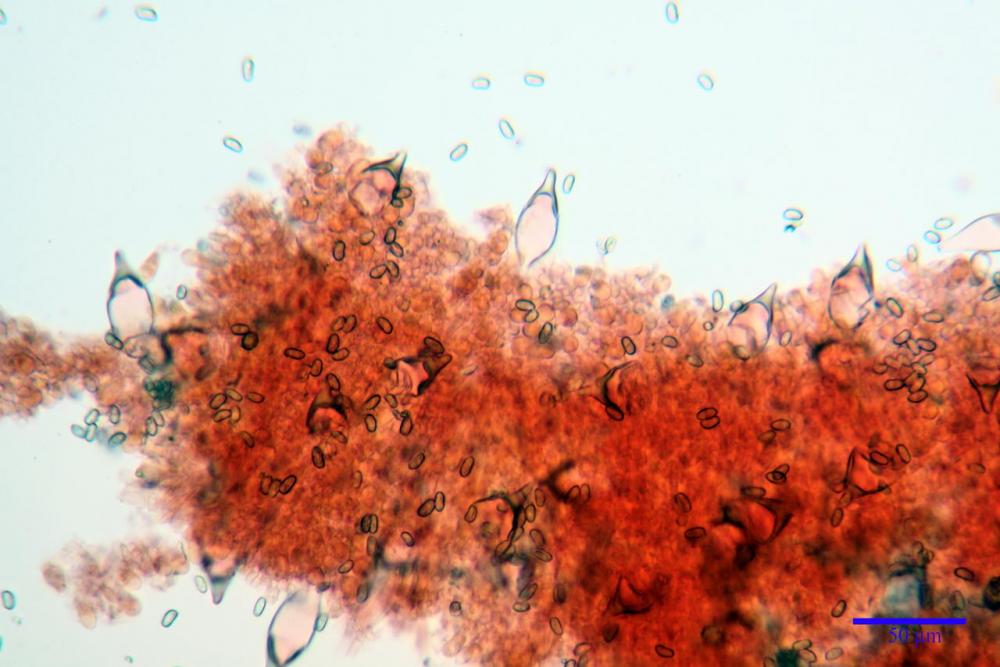
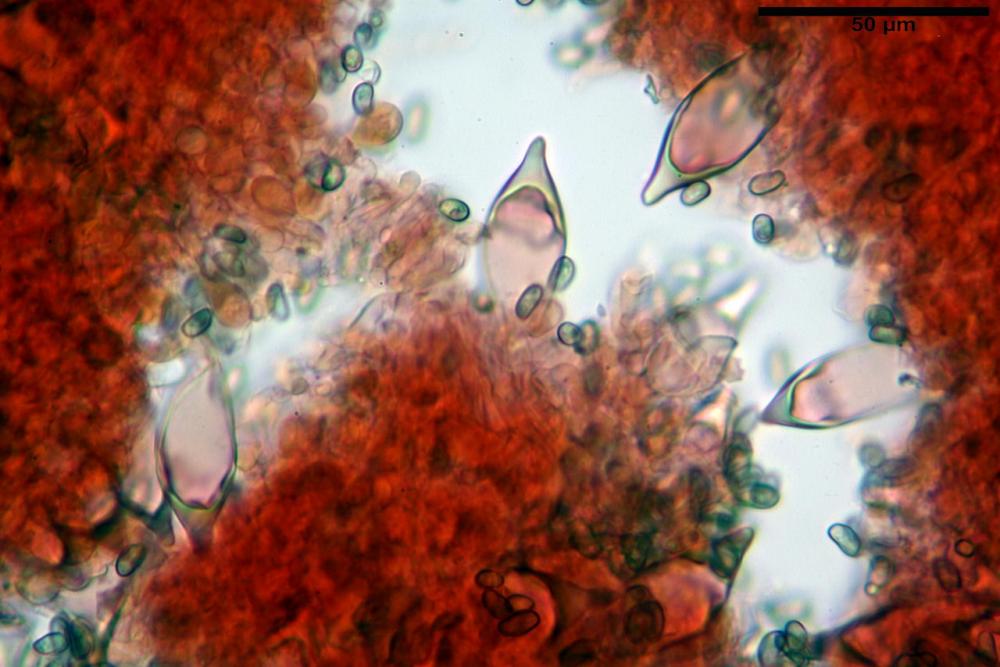
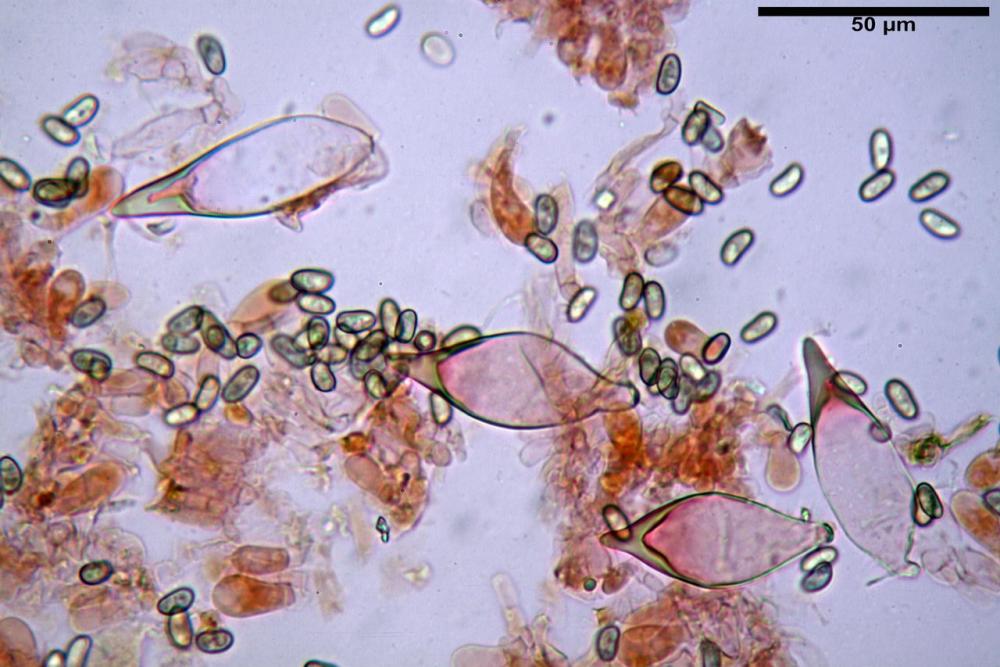
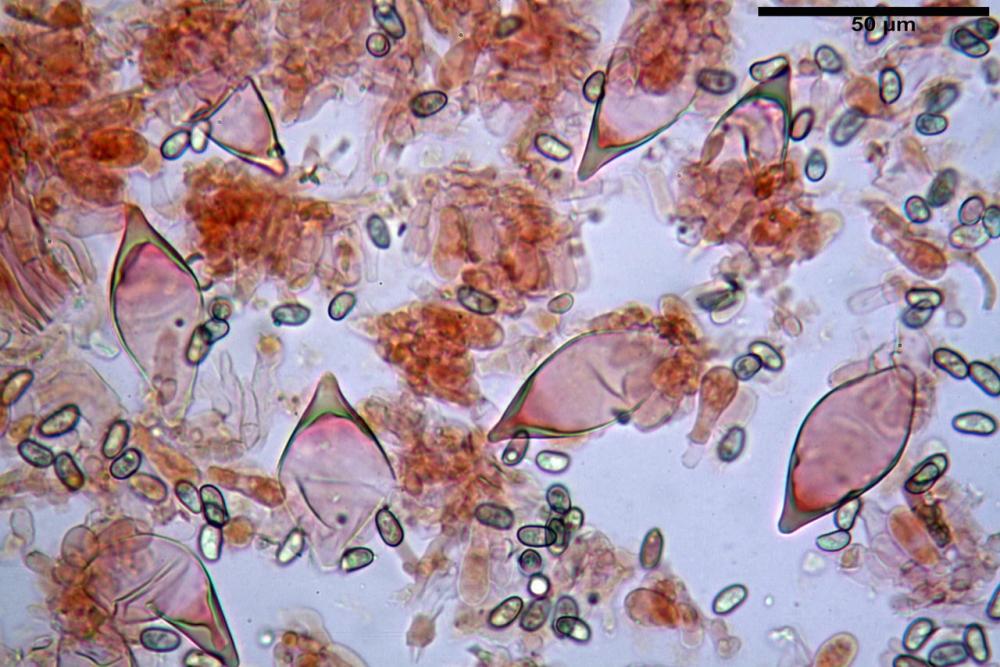
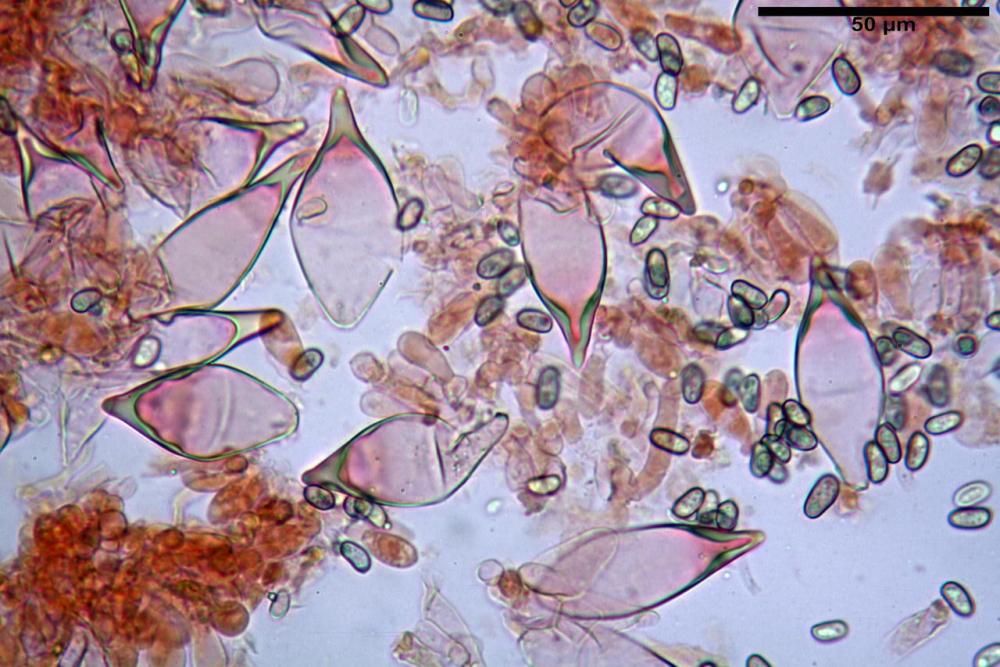
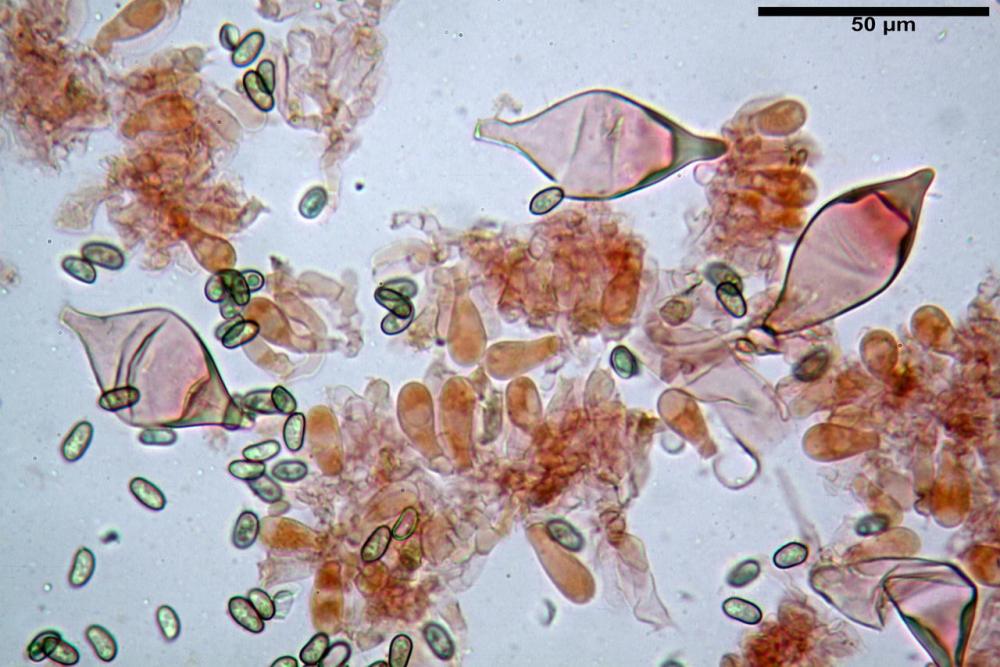
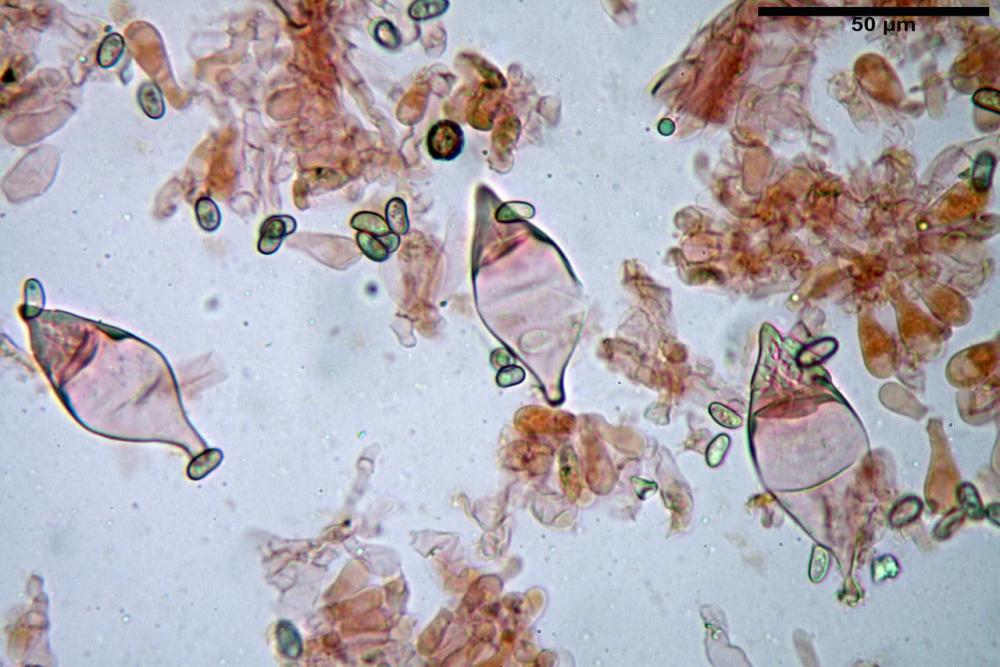
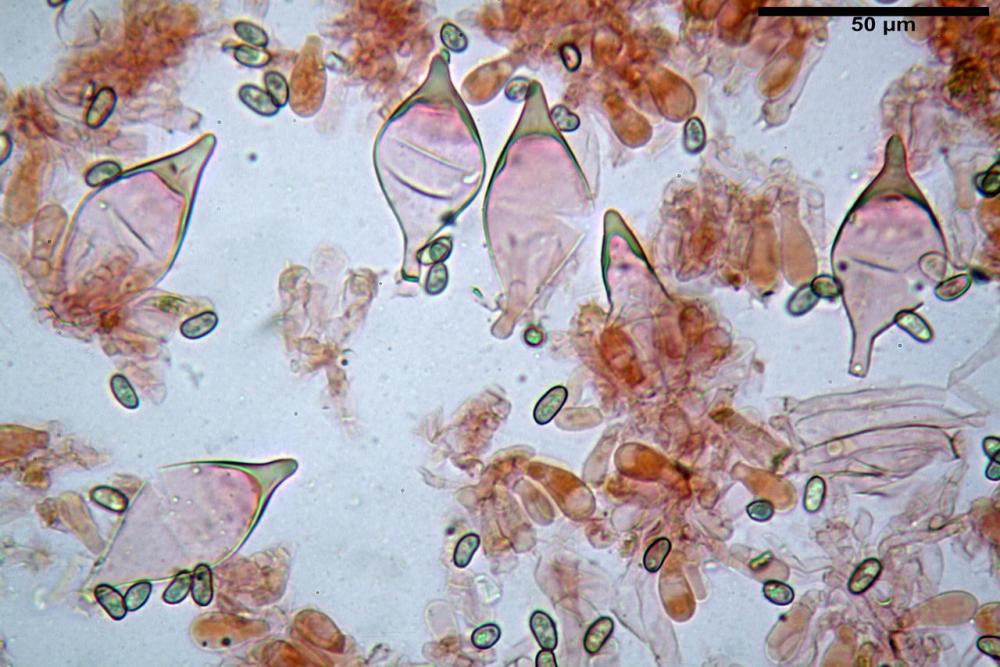
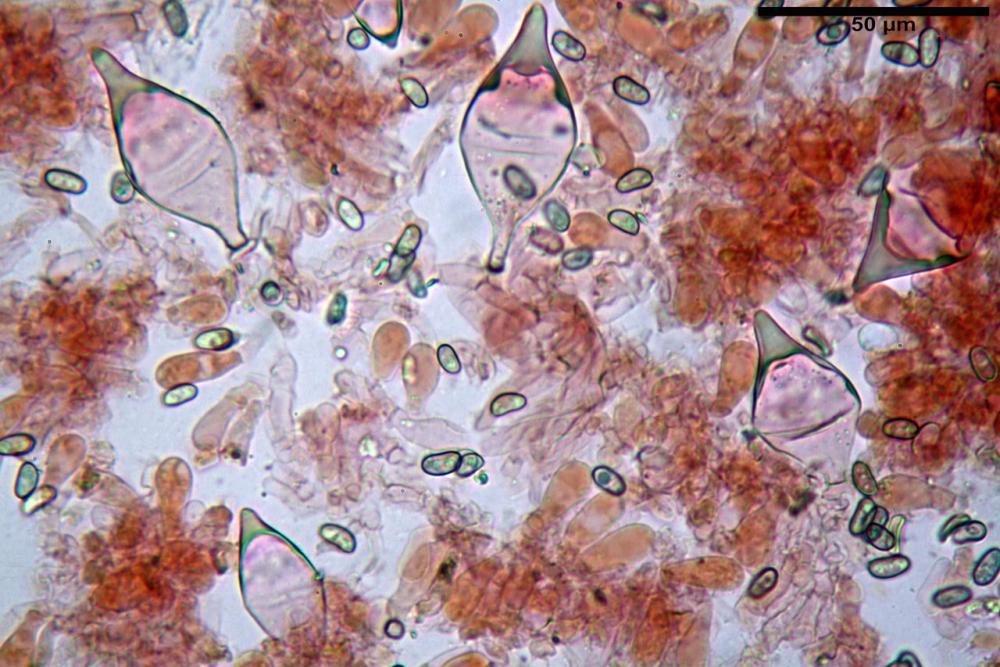
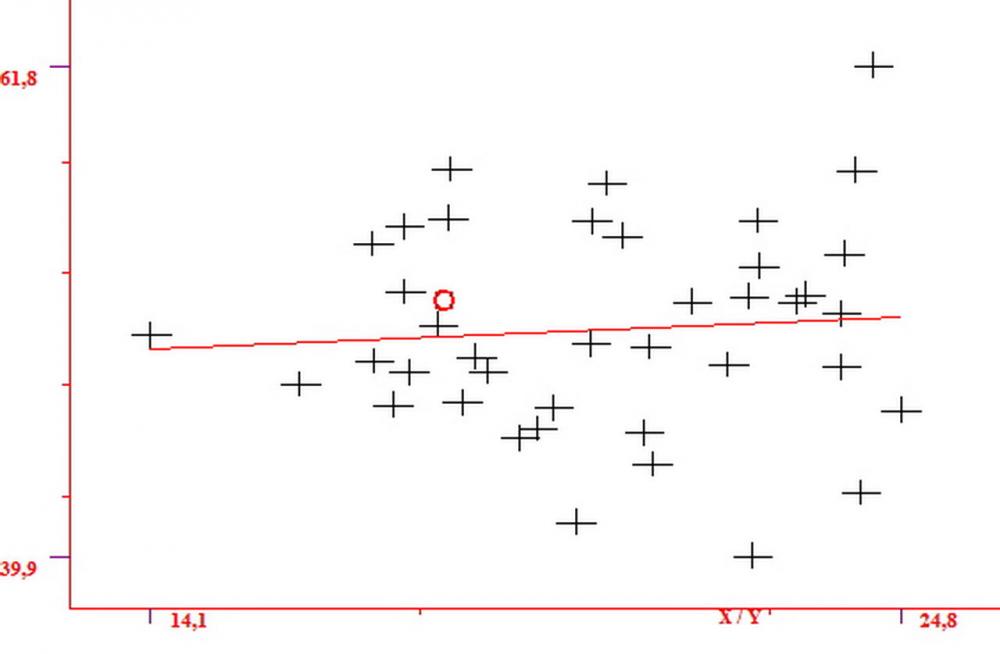
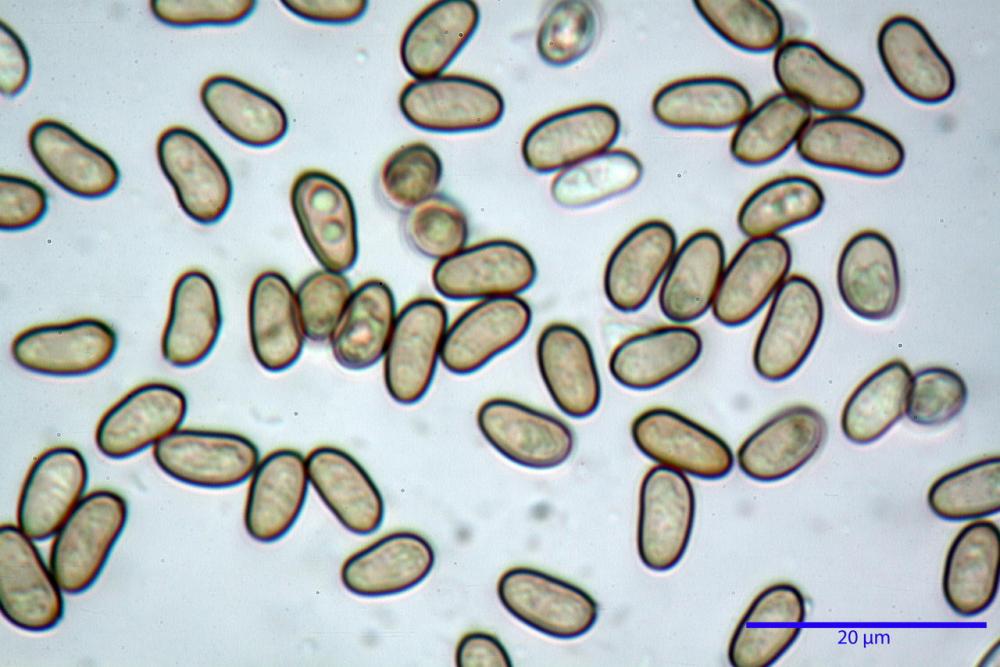
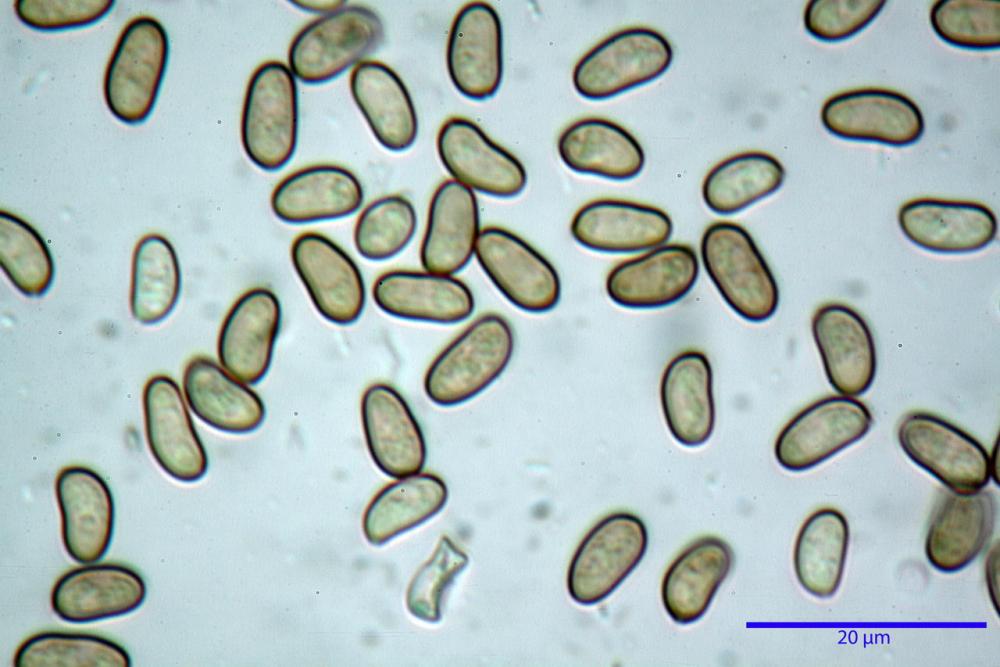
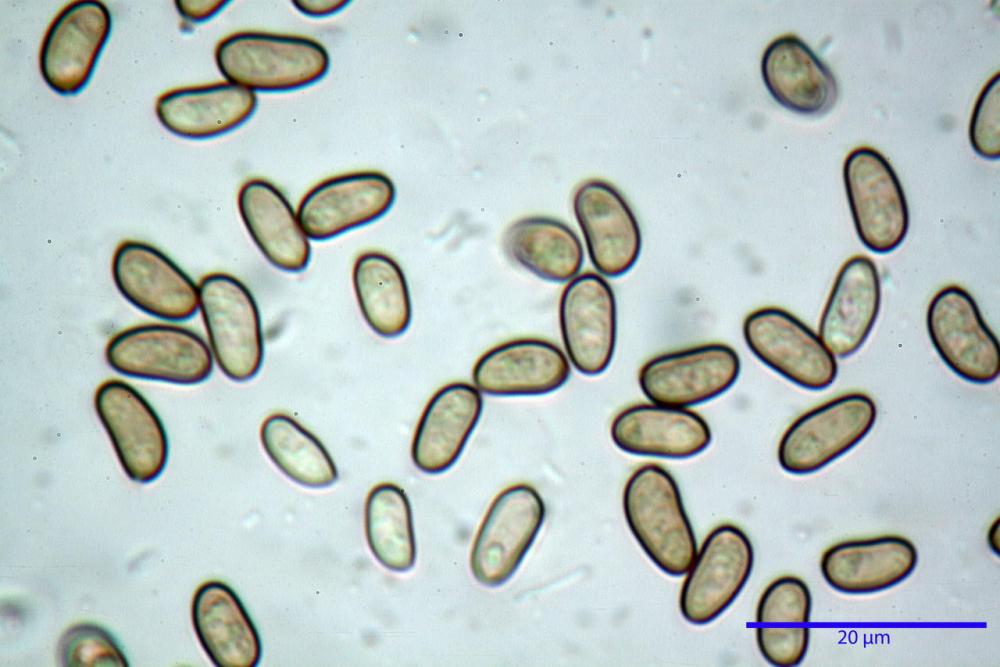
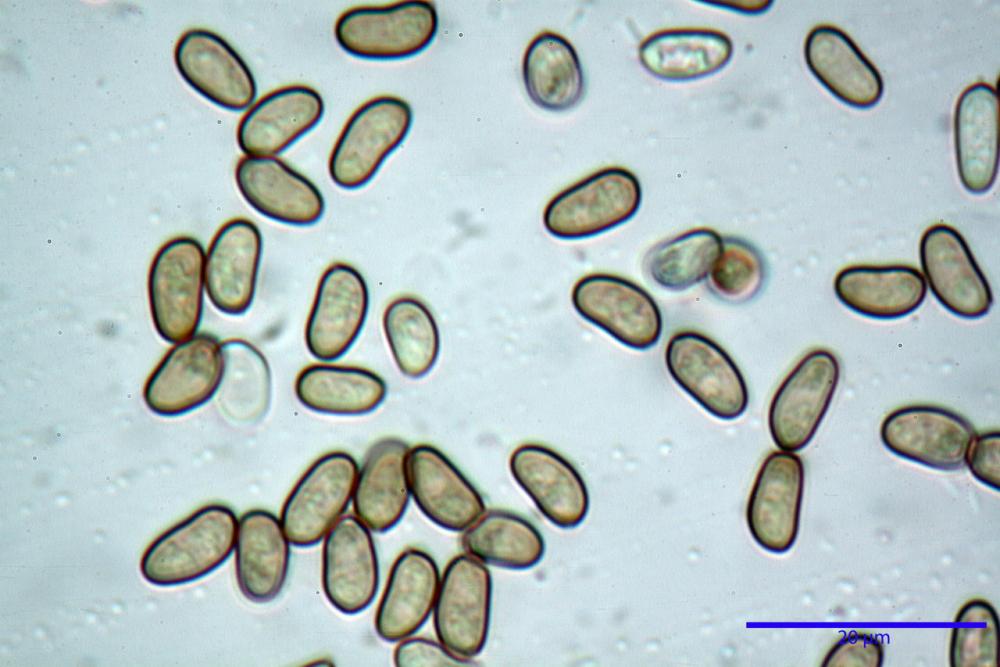
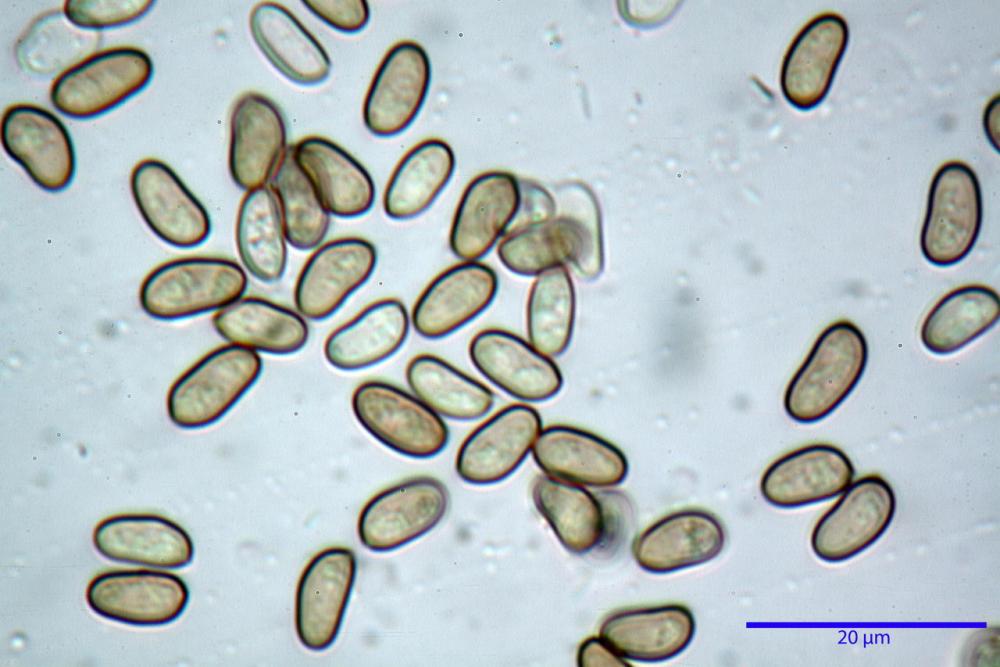
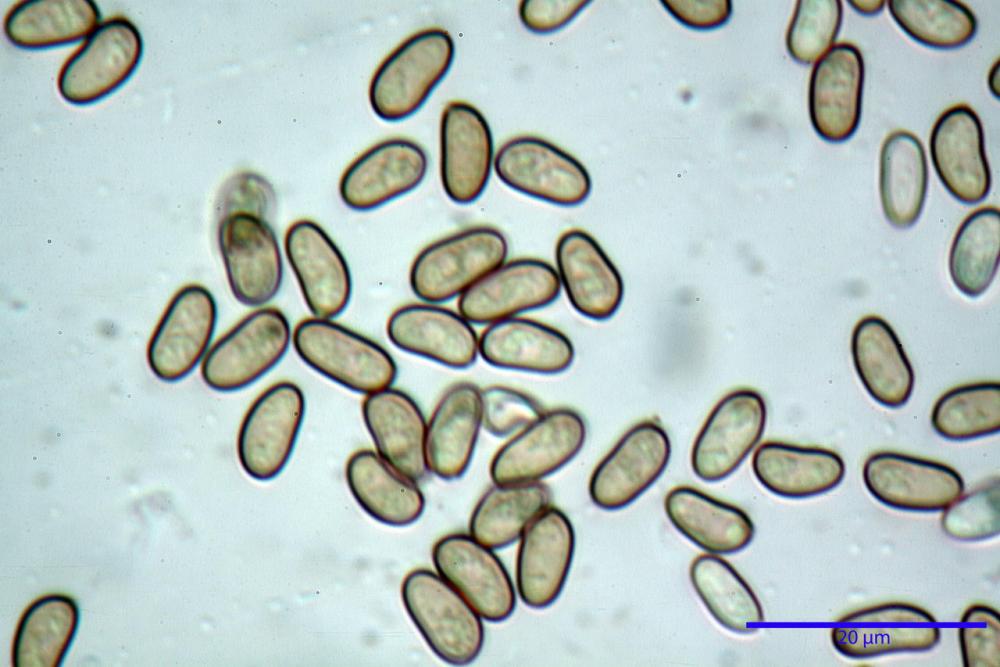
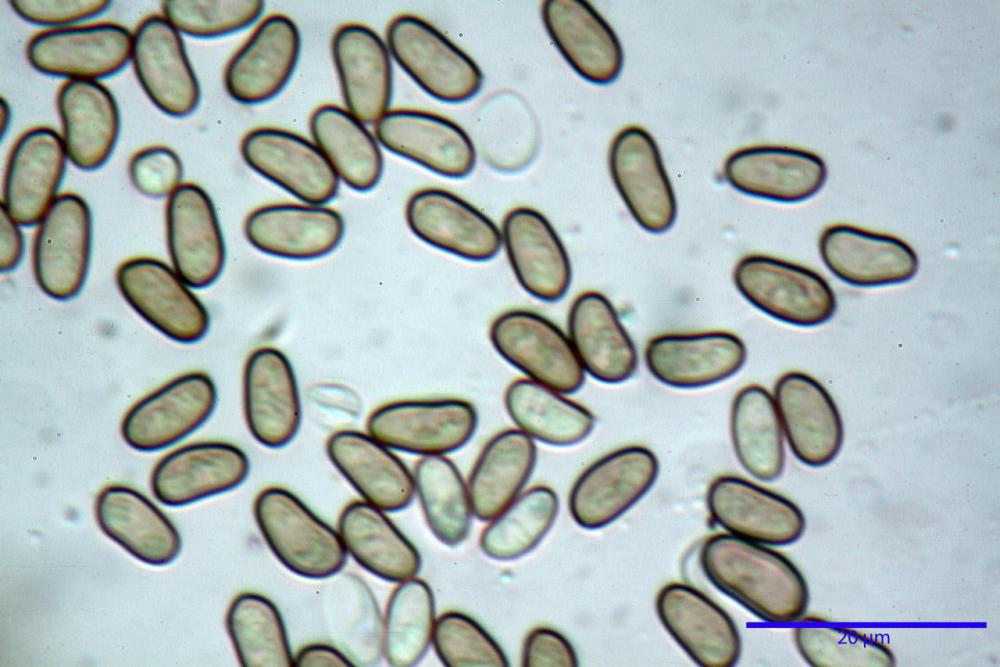
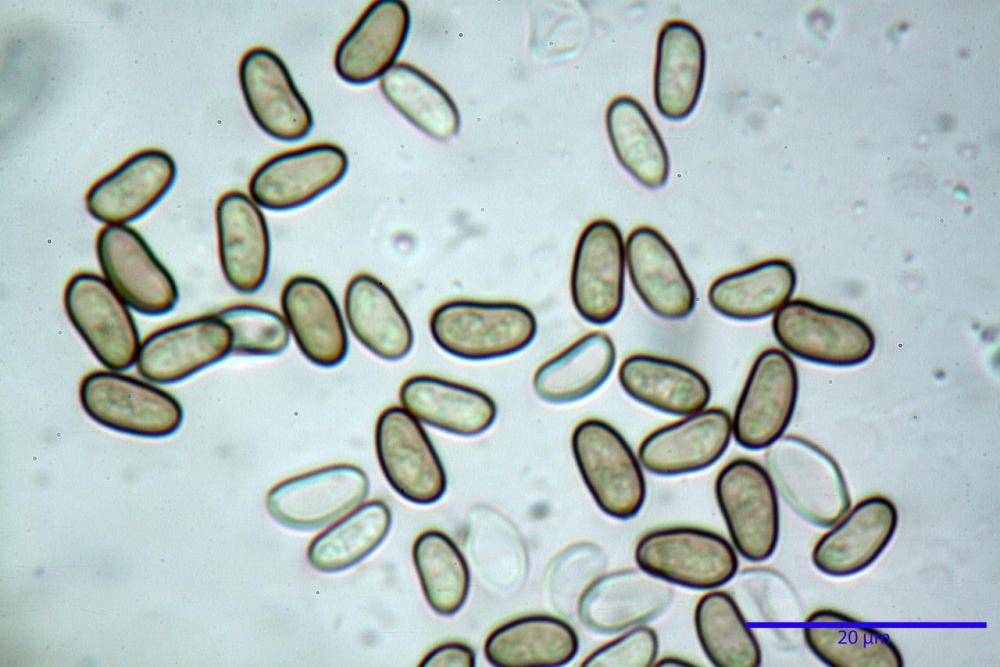
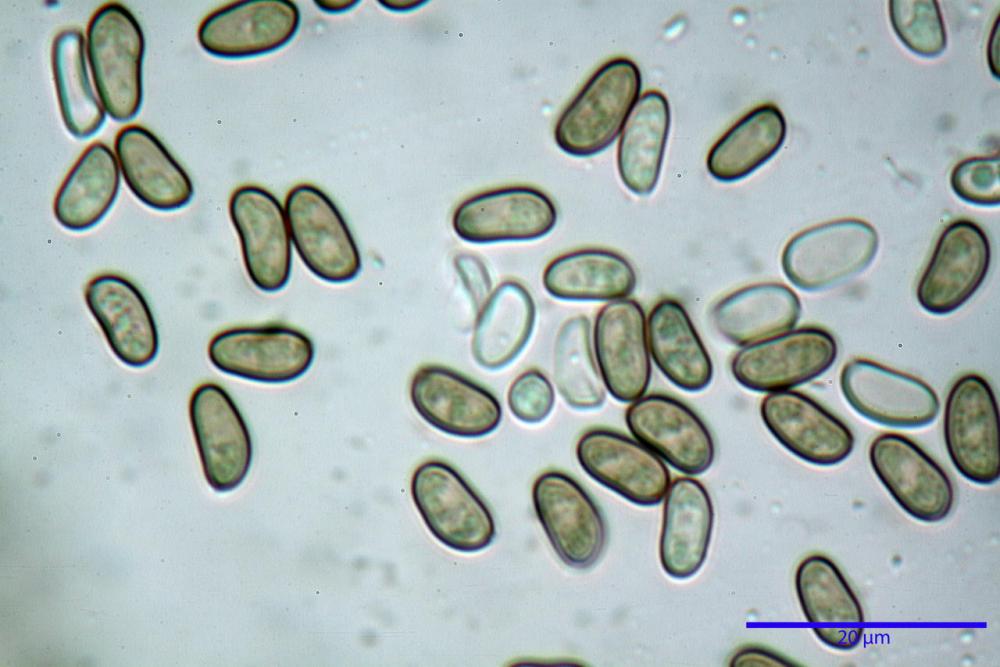
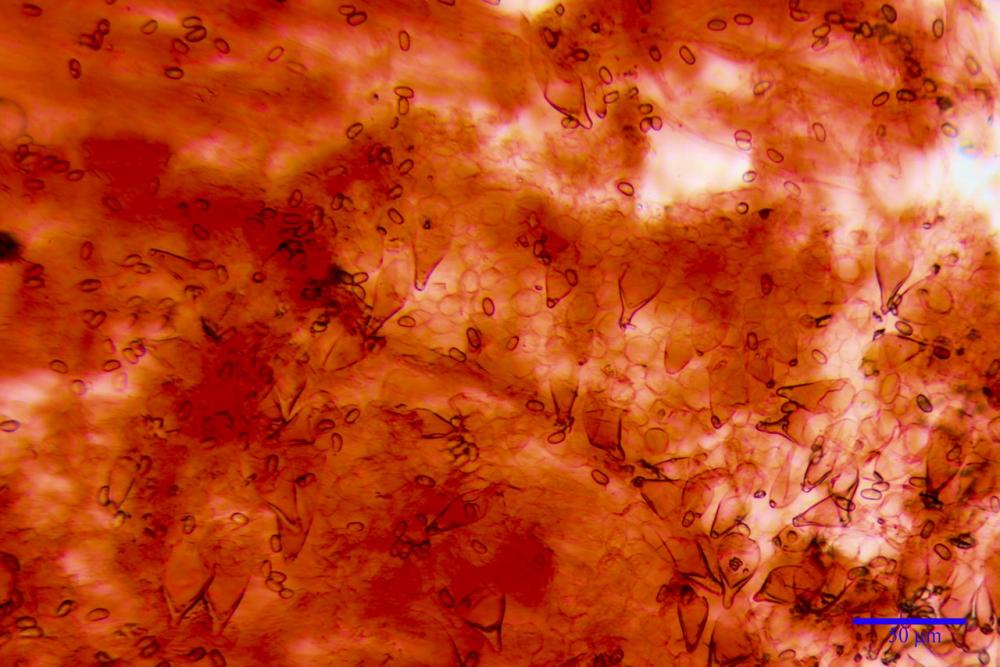
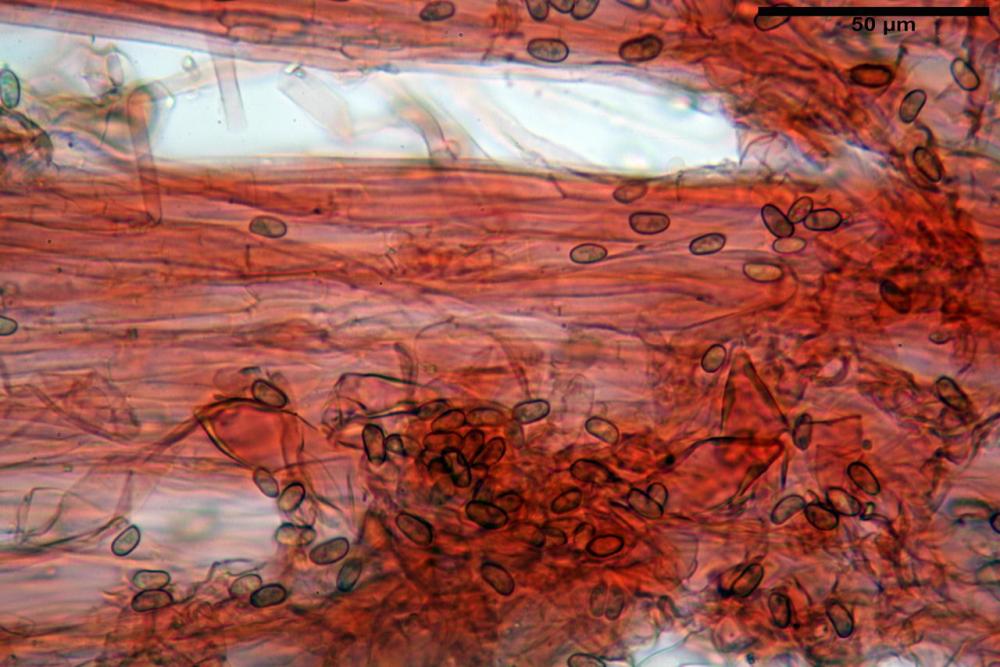
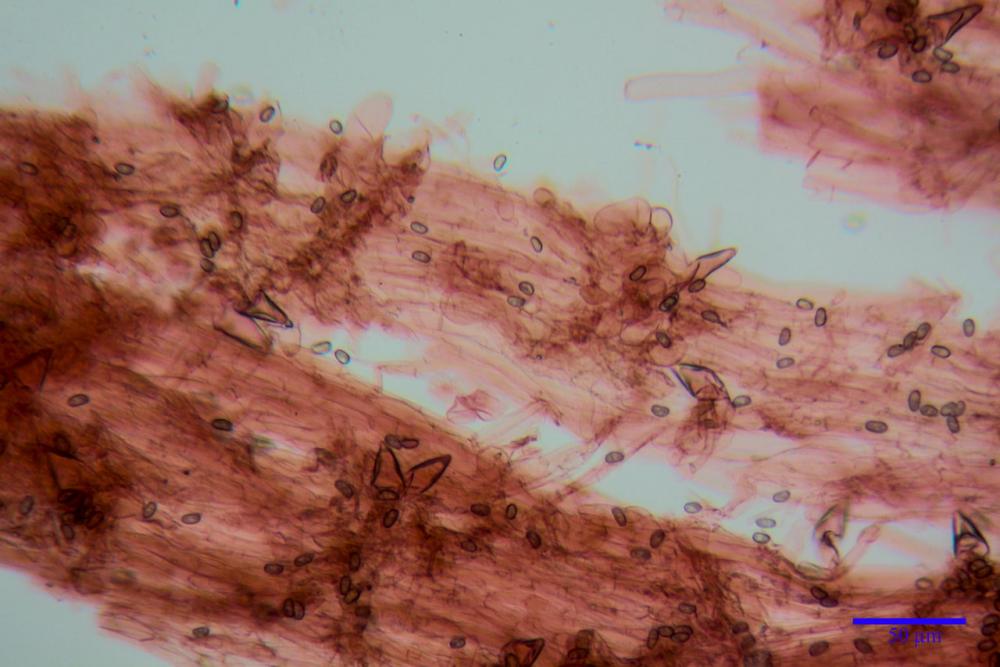
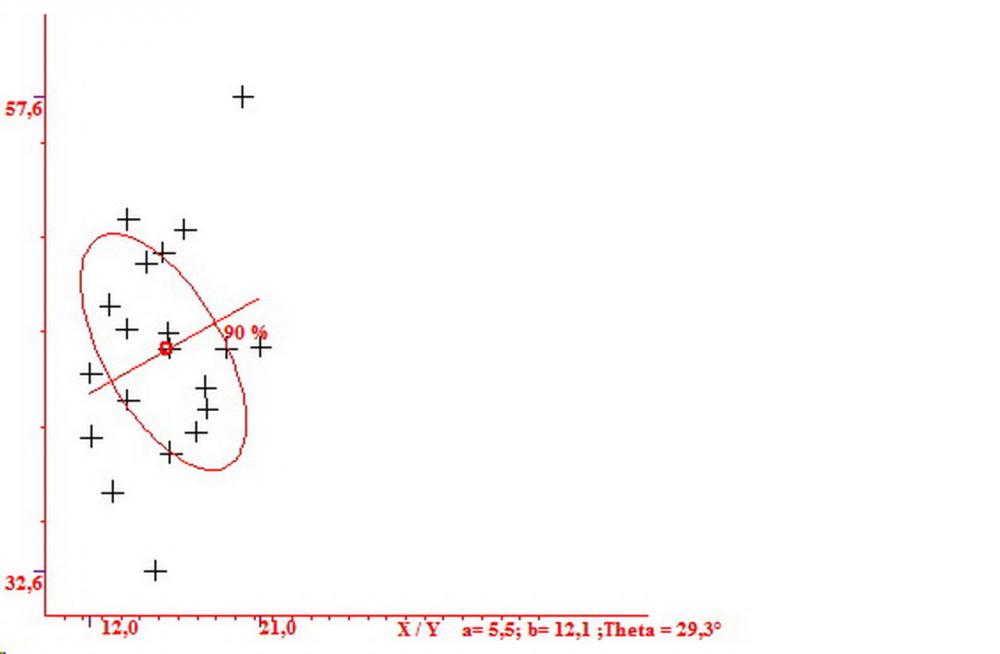
Spore (7,54)7,83-8,49-9,15(9,61) x (3,81)4,11-4,35-4,61(5,00) µm, Q. = (1,74)1,81 - 1,95 - 2,1(2,22); Vol. = (57,3)71,03 - 96,96(121,0), Vol.m.= 84,5 µm3, in proiezione laterale faseoliformi, di rado ellissoidi, in proiezione frontale subcilindriche, in rari casi ovoidi. Lisce, a contenuto granulare verdastro alle volte con una o due guttule centrali, ialine, in H2O molto chiare al microscopio ottico, parete spessa fino a 0,6 µm, prive di poro germinativo, appendice ilifera poco evidente.
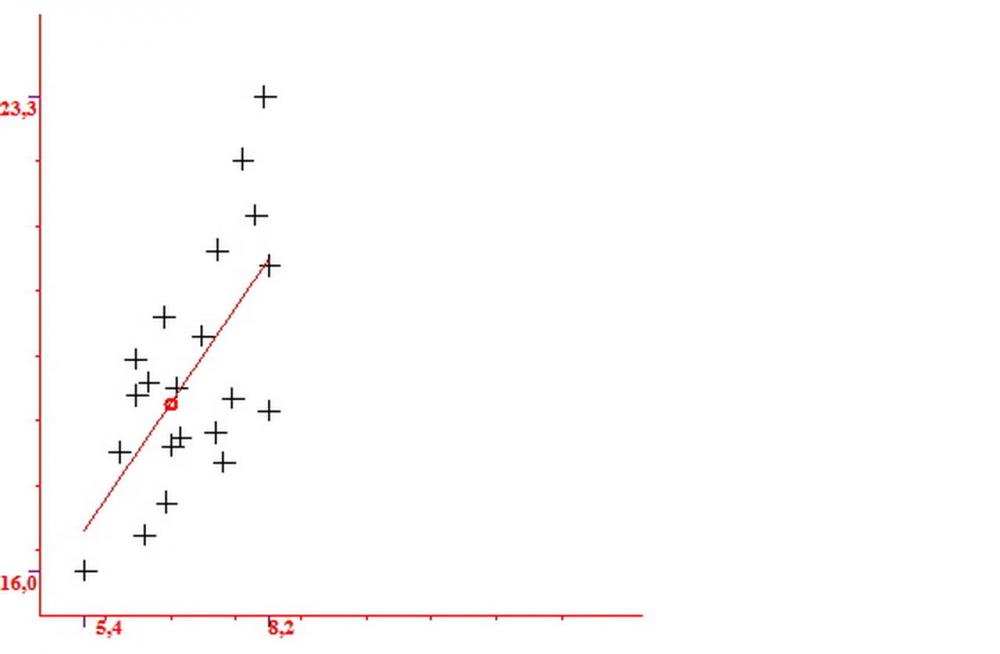
Basidi (16,0) 16,6 -19,0- 22,1 (23,3) x (5,4) 6,0 -7,0- 8,2 (8,24) µm, Q. = (2,2) 2,3 - 2,72 - 3,0 (3,1); Vol. = (246) 334 - 735 (814), Vol.m.= 510µm3, clavati, ialini, tetrasporici.
Trama lamellare (5,2) 5,9 - 13,9 (14,9) µm, costituita da ife inflate, ristrette ai setti, in NH4OH 2% ialine nella porzione centrale, lieve pigmentate negli strati esterni, giallo-brunastre nel subimenio.
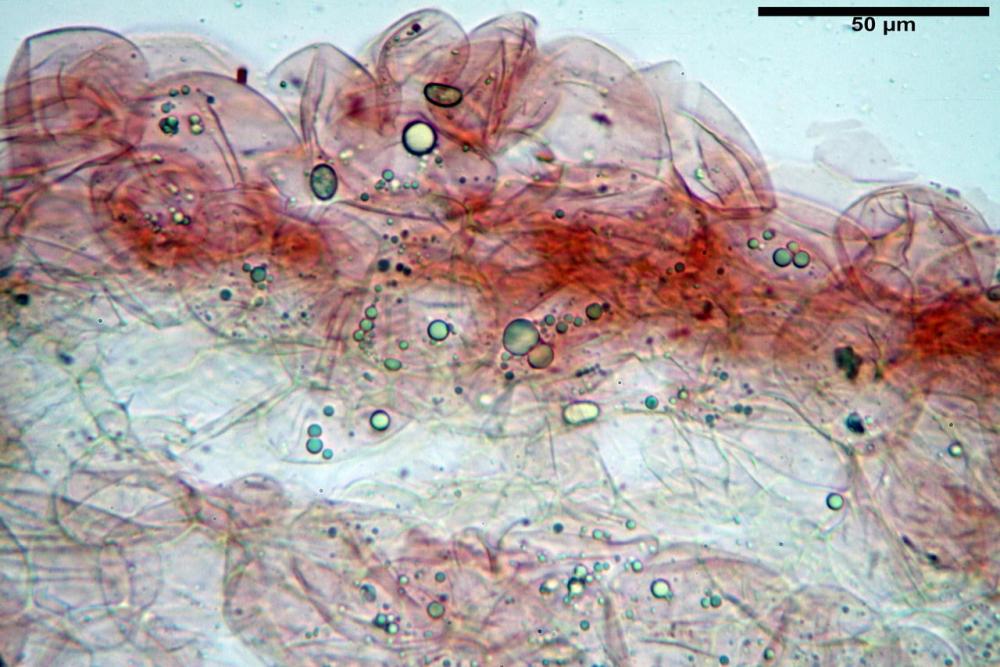
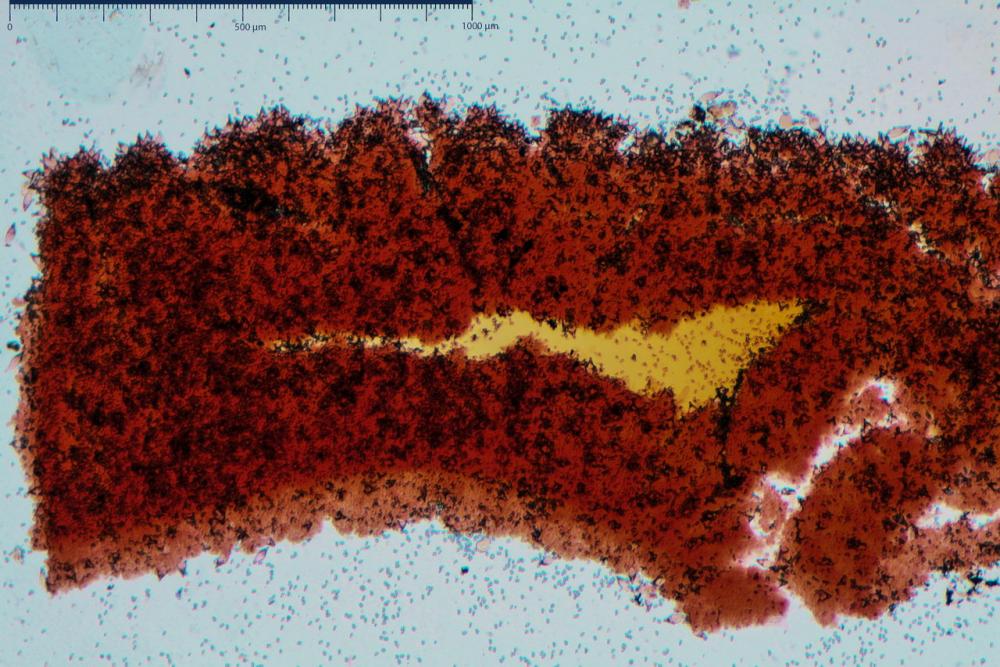
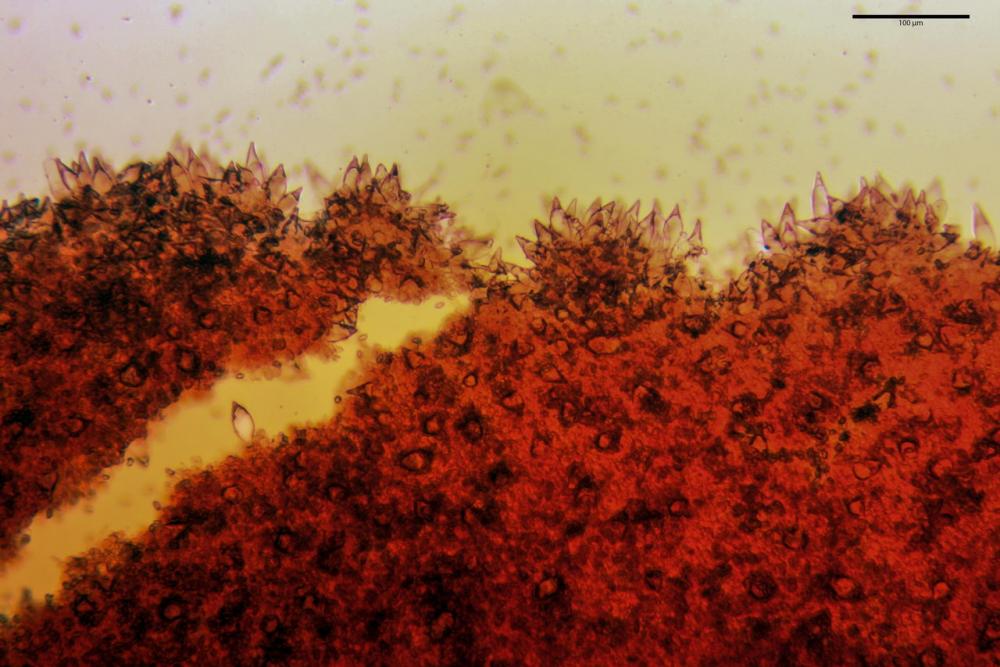
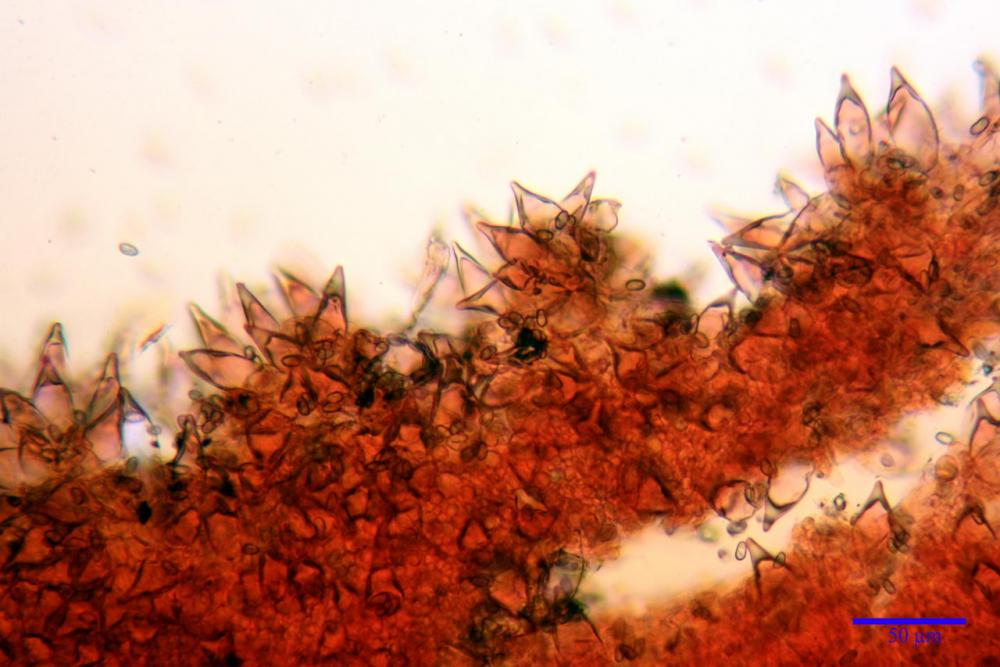
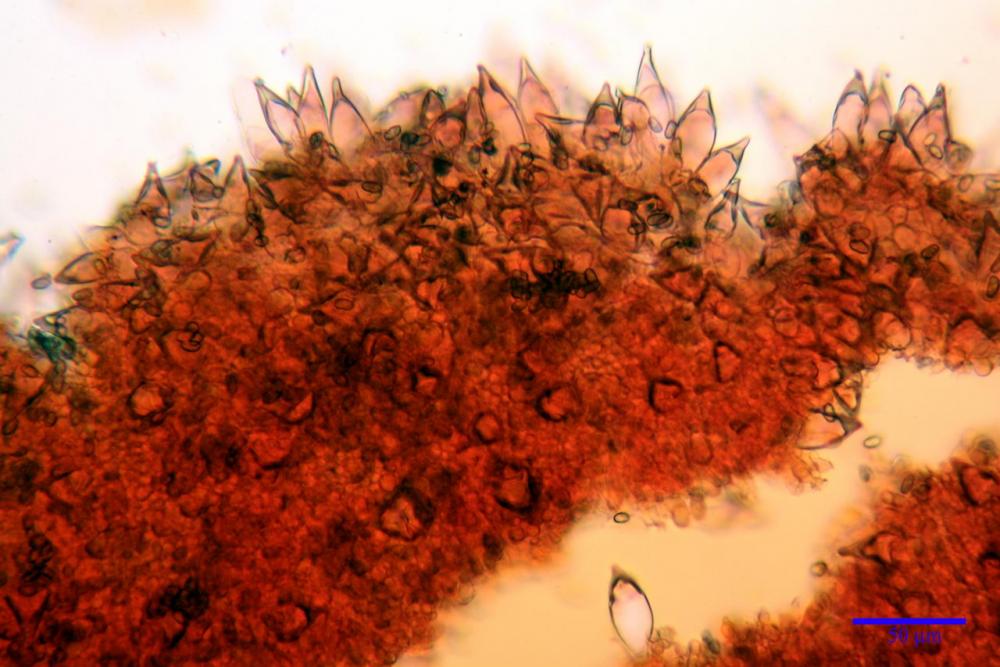
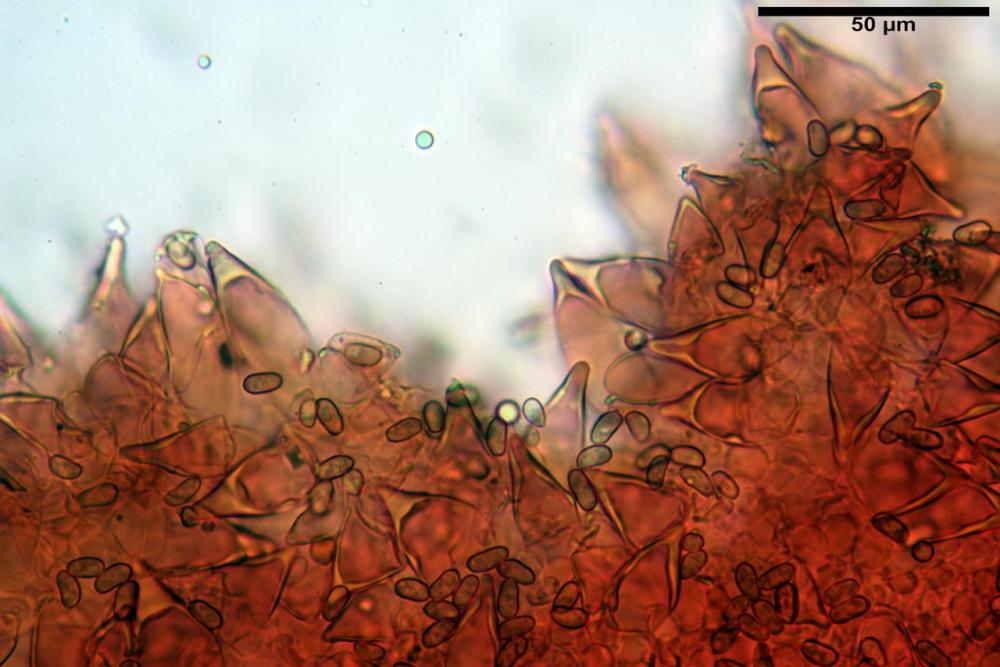
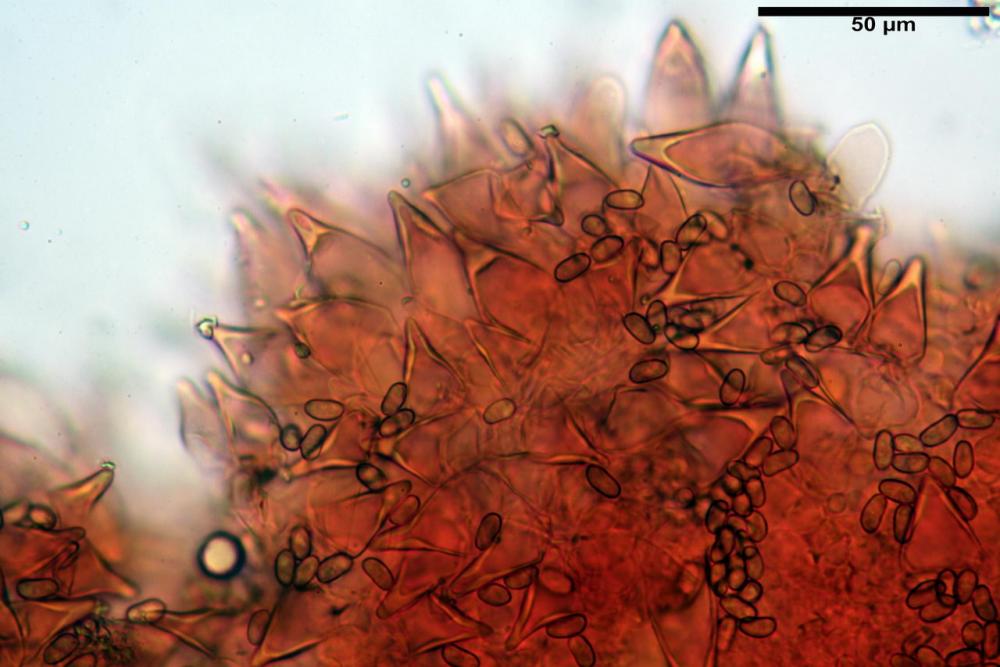
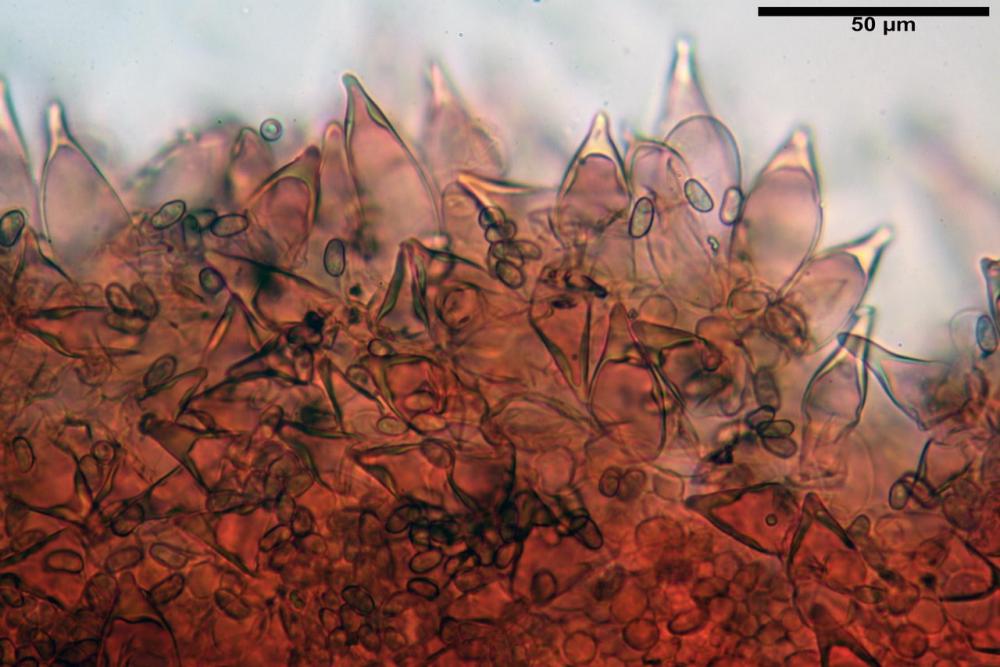
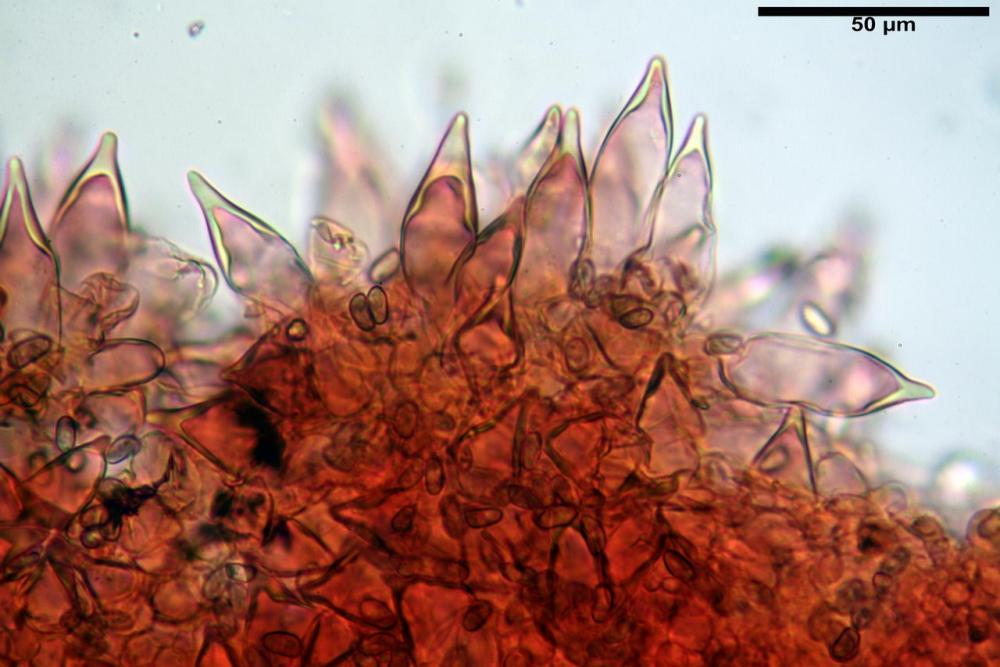
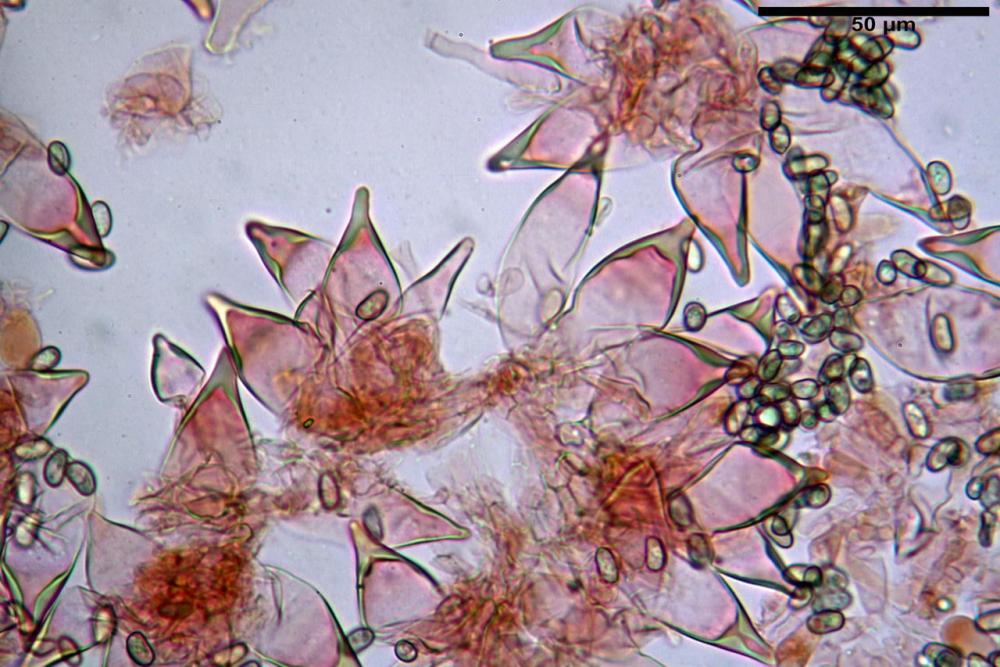
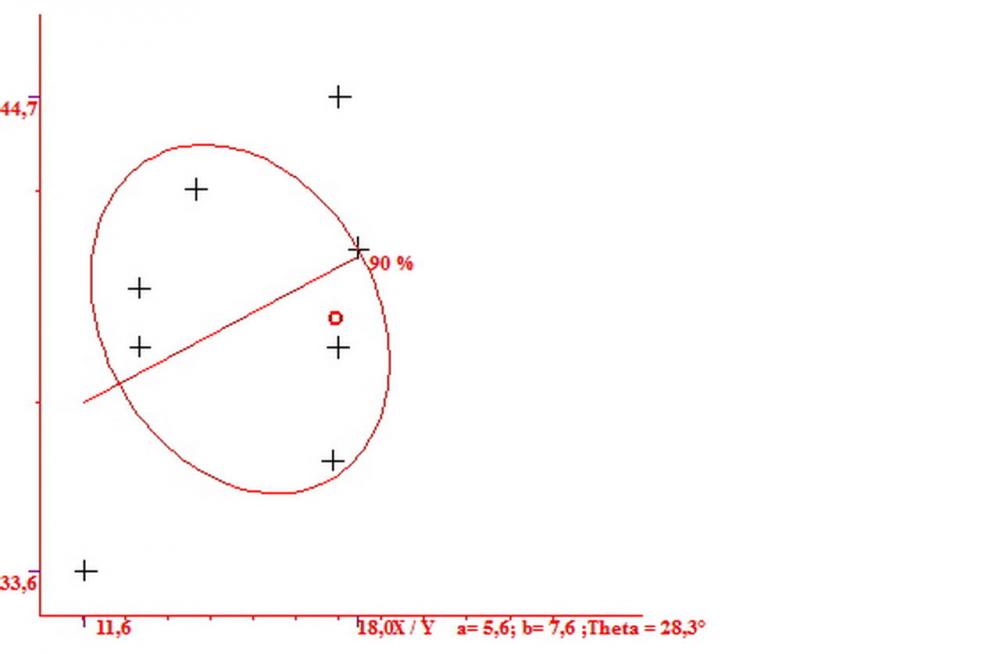
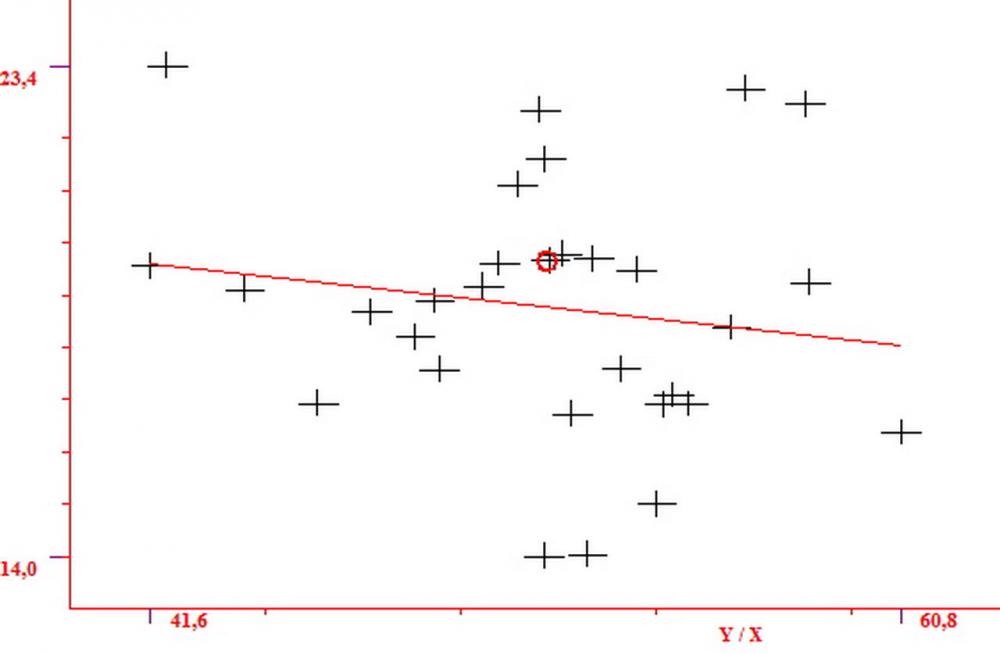
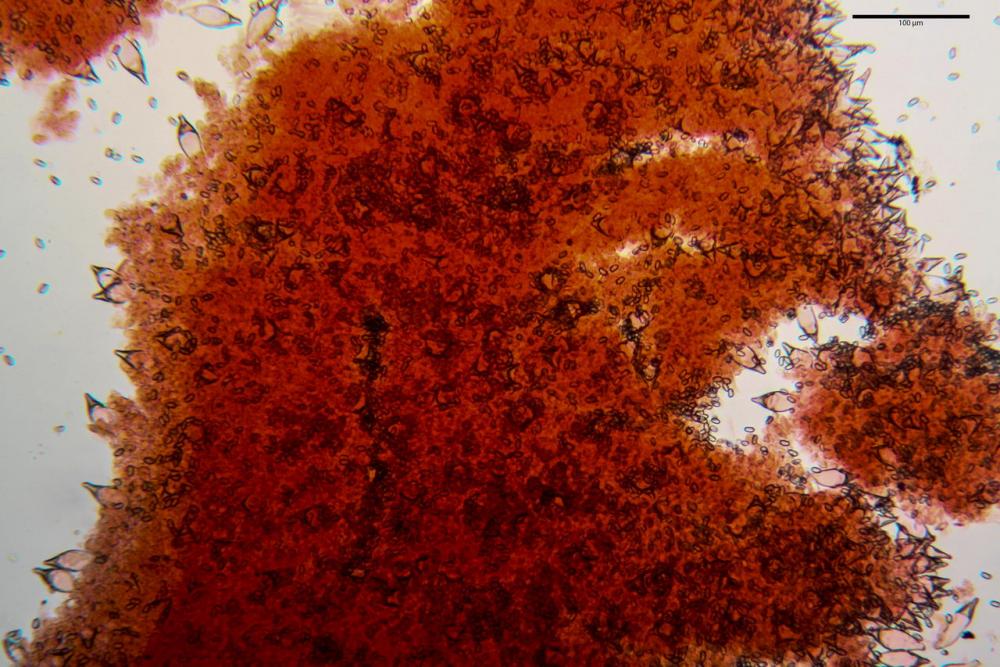
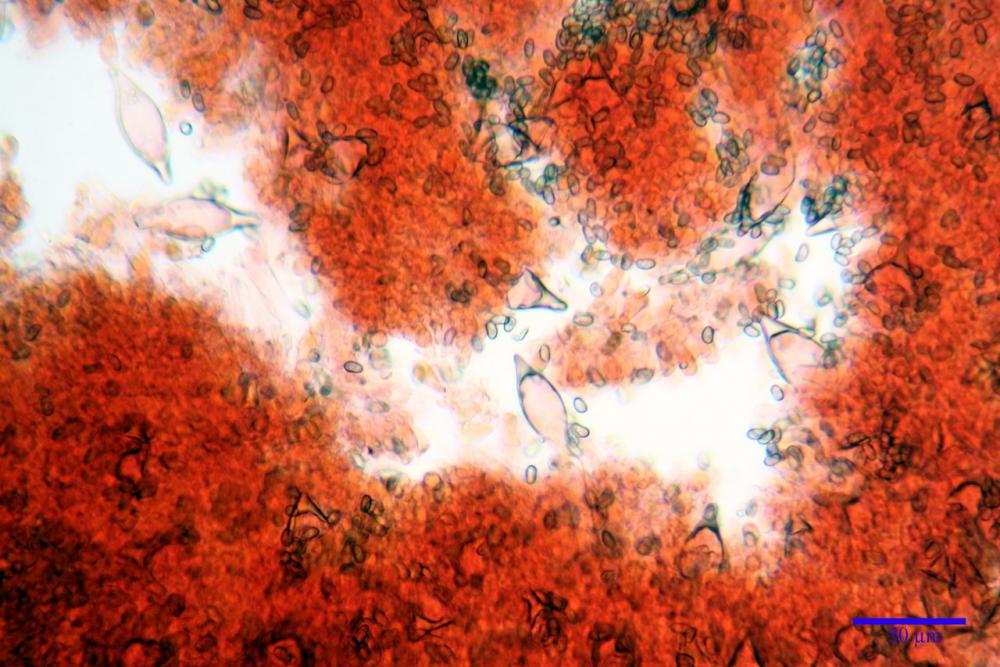
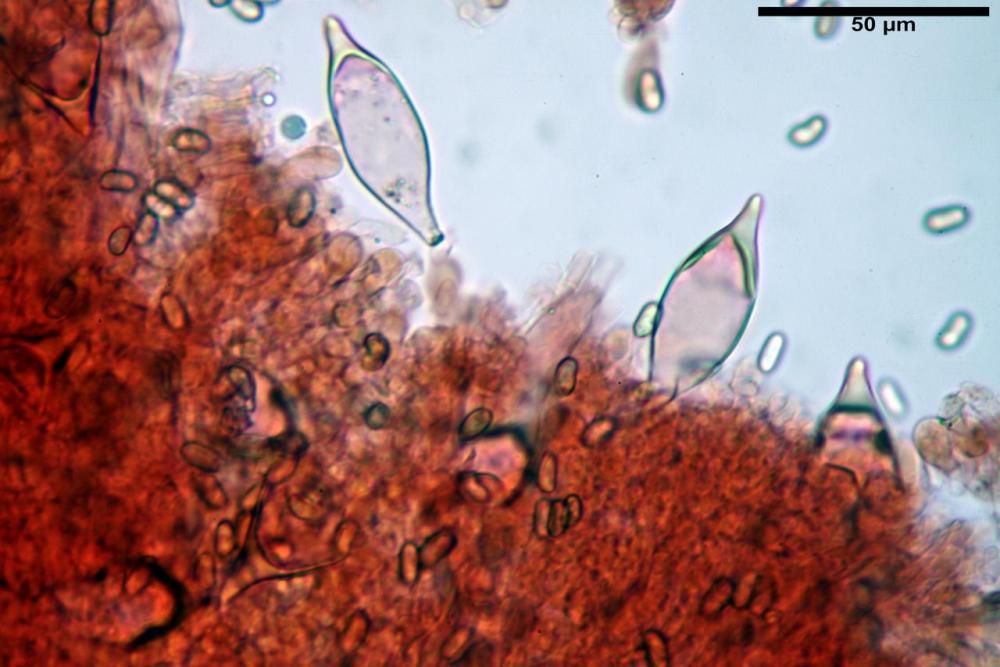
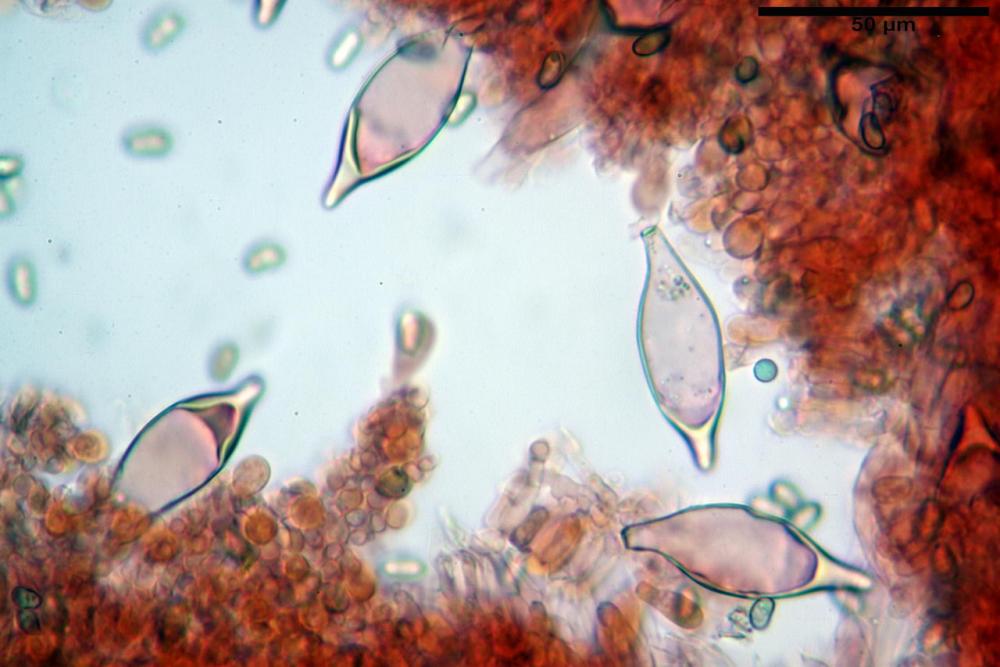
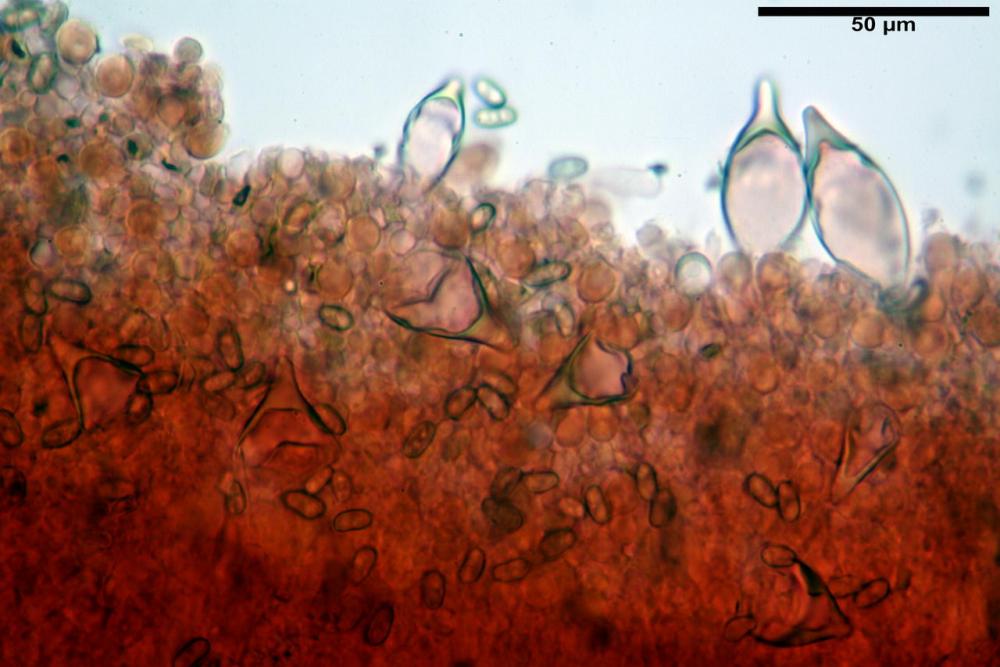
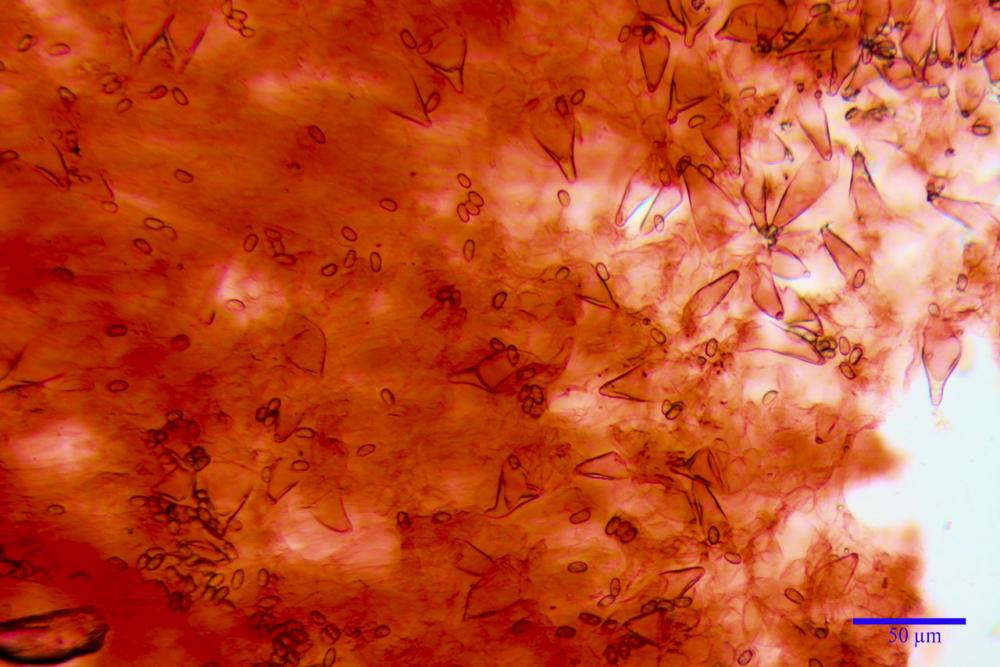
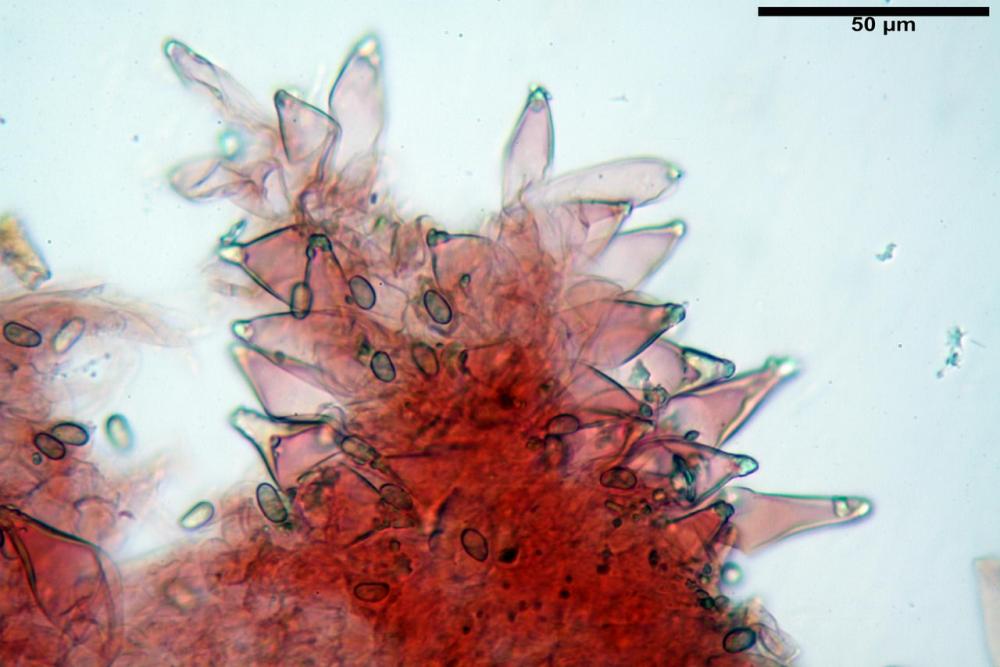
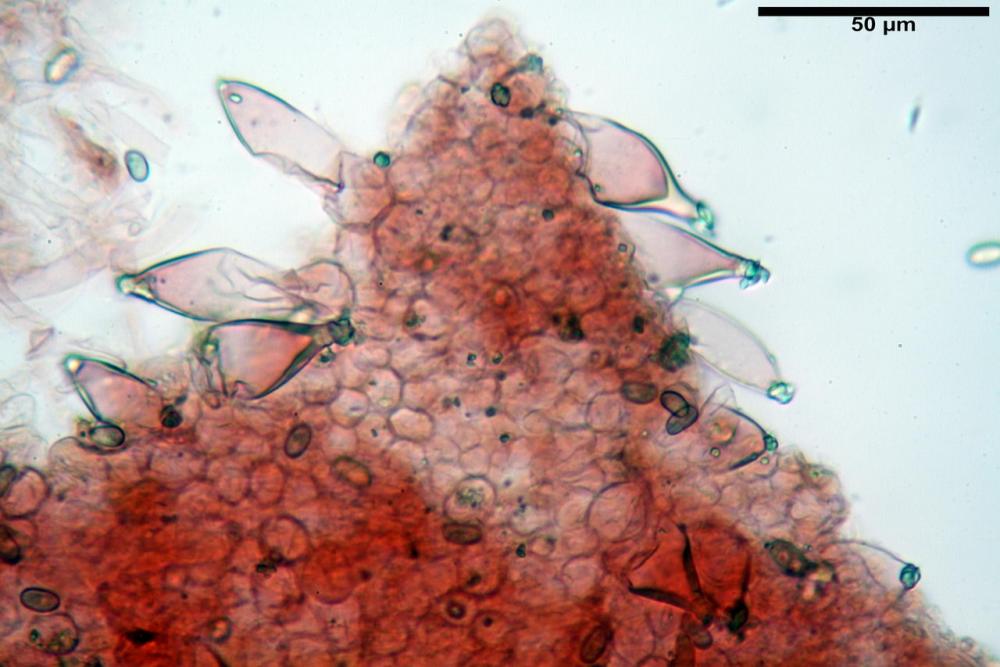
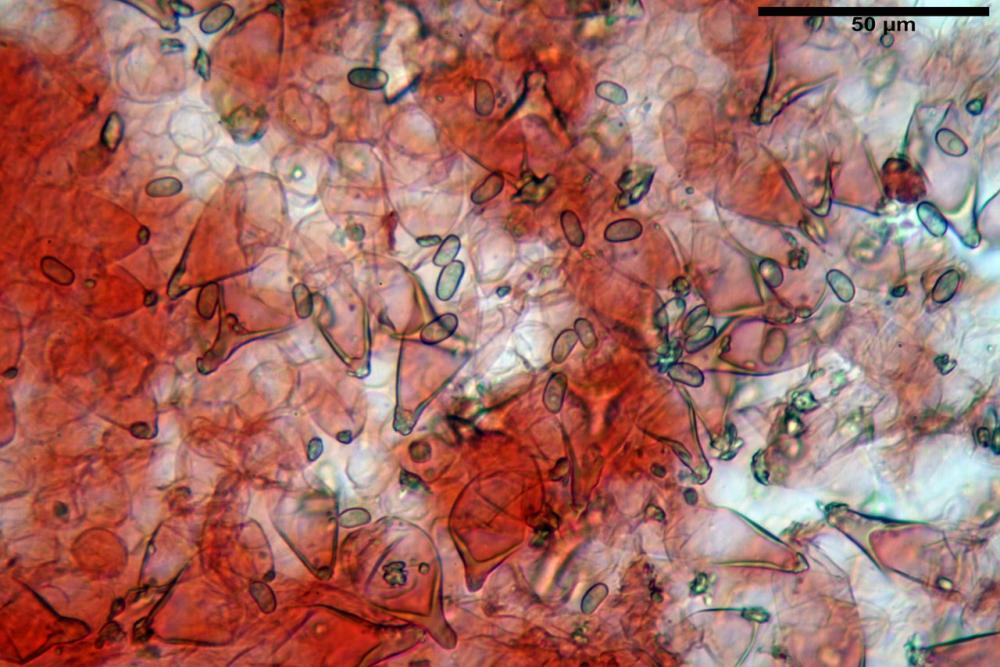
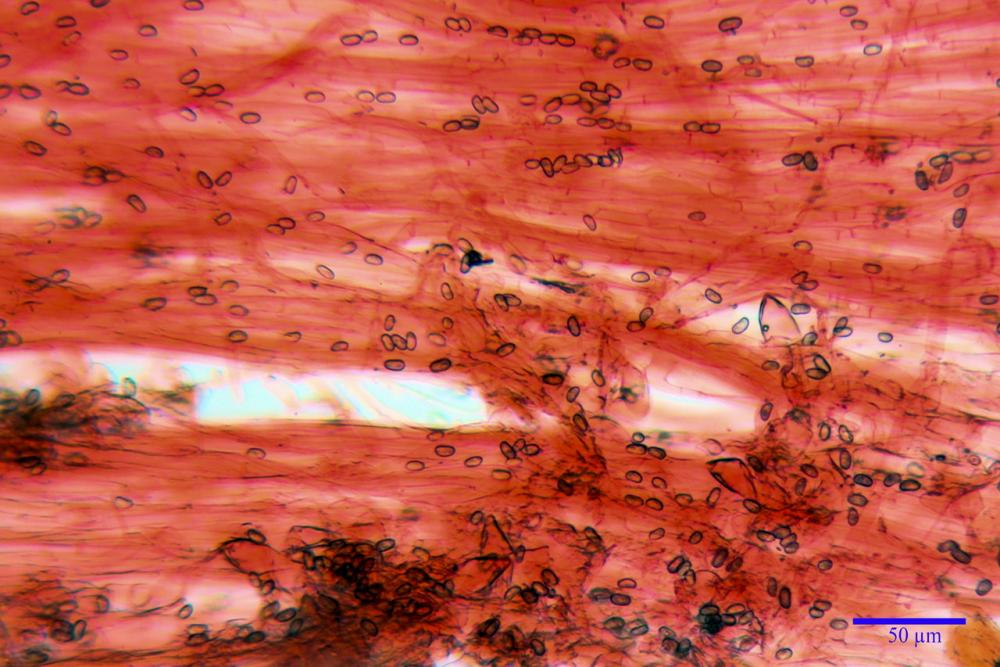
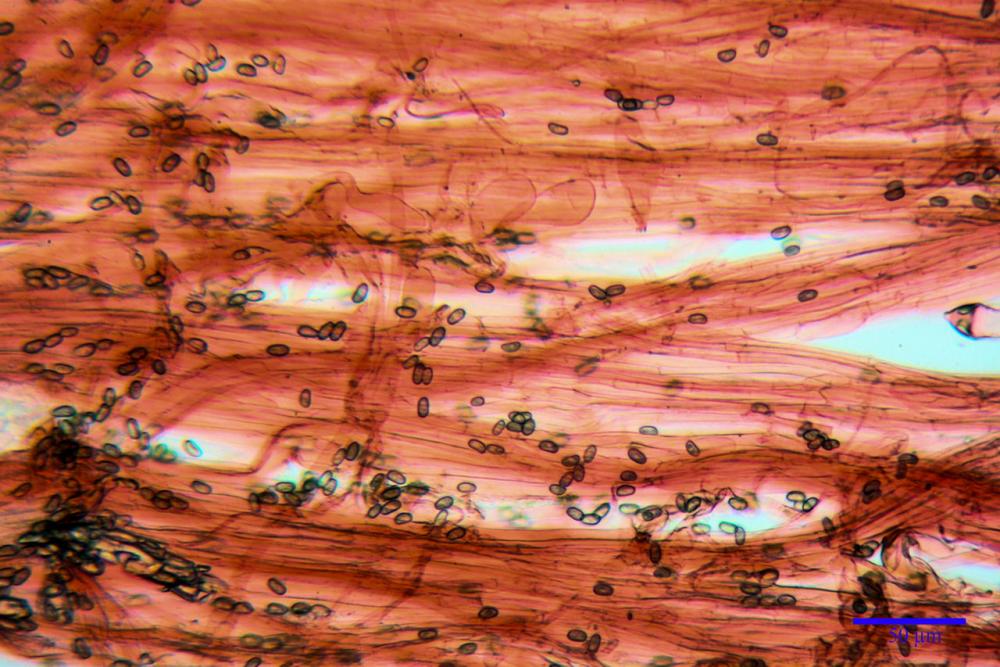
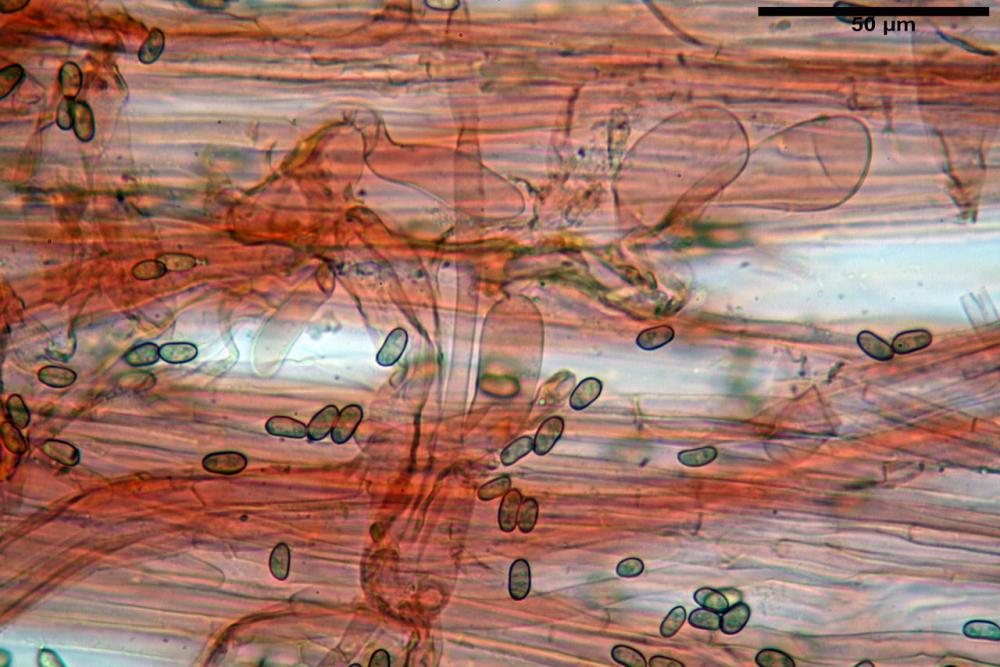
Cheilocistidi metuloidi (41,6) 44,1 -51,8- 58,2 (60,8) x (14,0) 15,1 -18,7- 22,6 (23,4) µm, Vol.m.= 9695 µm3, da strettamente a largamente fusoidi con apice acuto e corto pedicello basale, in alcuni casi assente, spessore parietale misurato al centro (0,58)0,76 - 0,99 - 1,45 (1,69) µm, all’apice (2,16) 2,26 - 2,55 - 3,12 (3,74) µm. Muricazione molto scarsa o quasi assente. In numero numerosissimo, tanto da rendere il filo lamellare completamente sterile.
Cheiloleptocistidi (33,5) 36,2 -39,5 - 42,8 (44,7) x (11,5) 12,7 -15,24 - 17,7 (17,9) µm, Vol.m.= 4976 µm3, clavati o subcilindrici, a parete sottile, ialini, da scarsi a molto rari, inframezzati ai metuloidi.
Pleurocistidi metuloidi (39,8) 44,1 -50,0- 56,3 (61,7) x (14,1) 17,3 -20,5- 24,1 (24,8) µm, Vol.m.= 11310 µm3 largamente fusoidi, con apice acuto e pedicello basale più o meno lungo, metuloidi, parete spessa fino a 3 µm, nella porzione apicale. Muricazione scarsa. Abbastanza numerosi, sparsi nella trama. Pleuroleptocistidi non osservati.
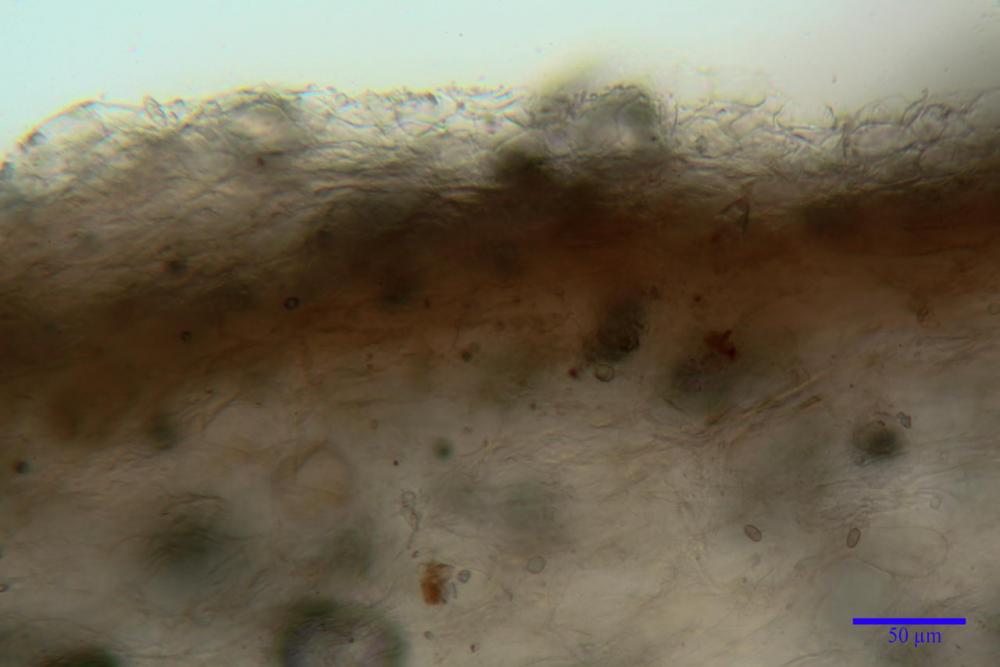
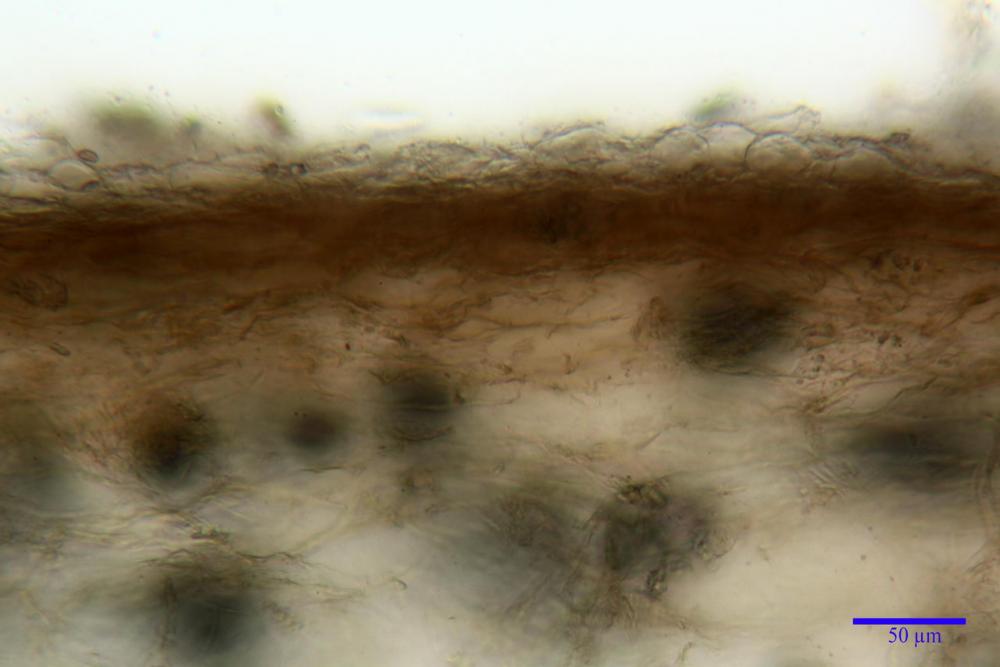
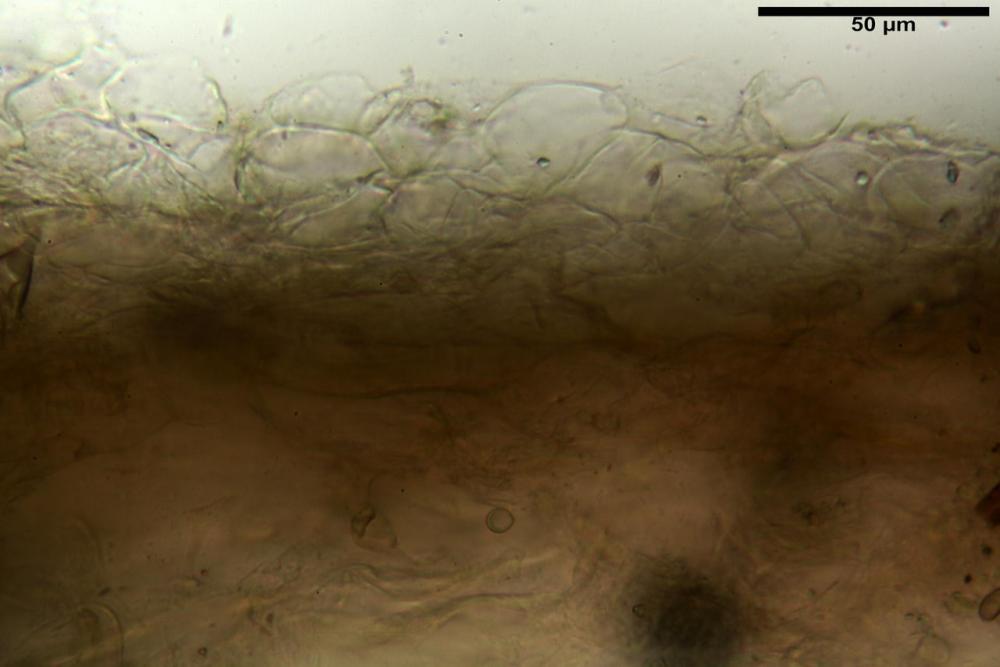
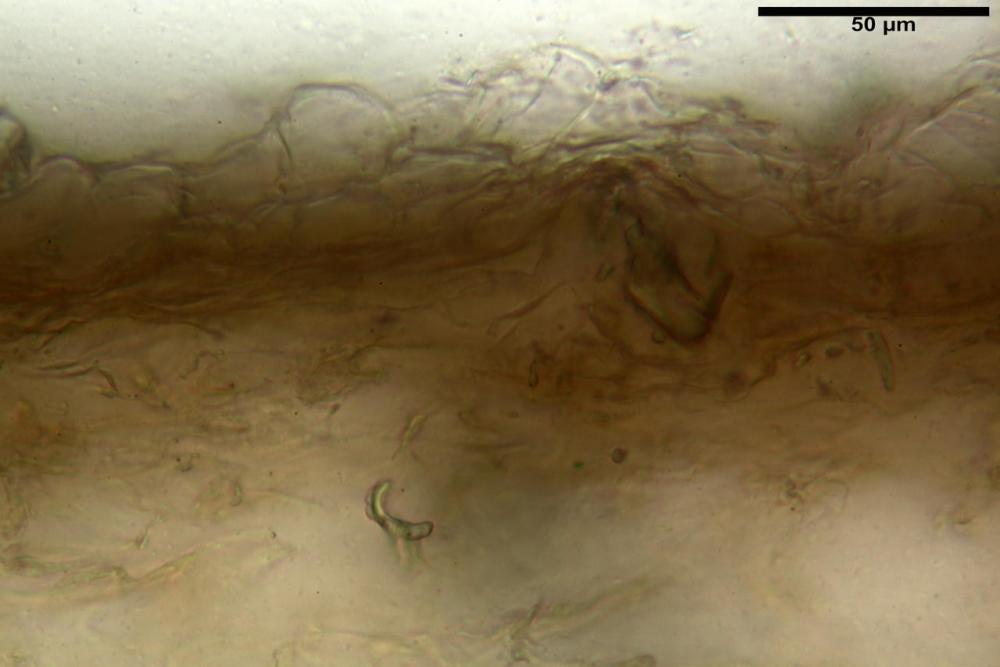
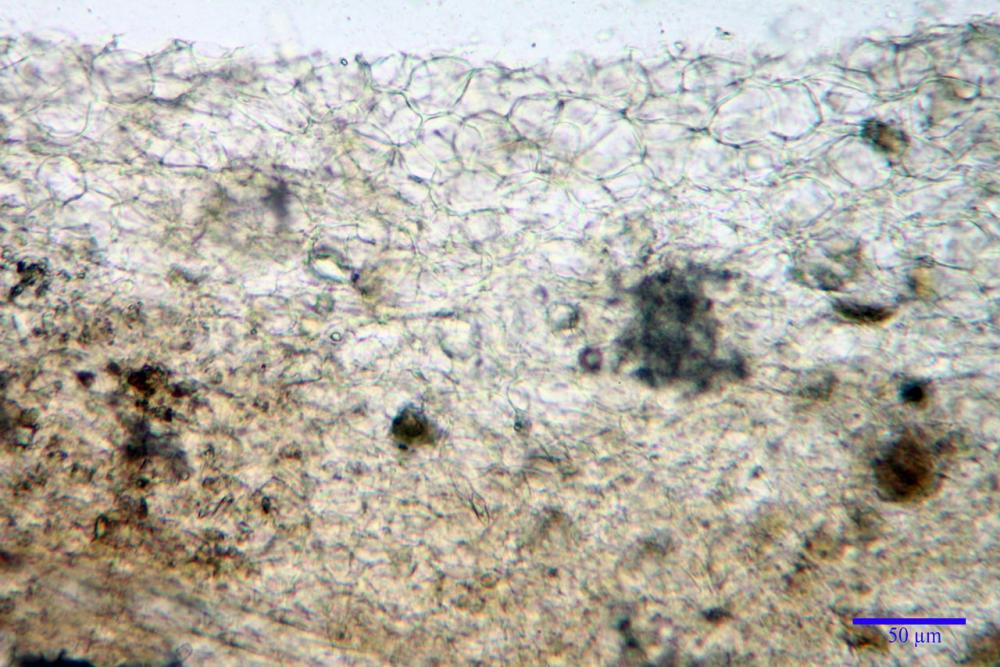
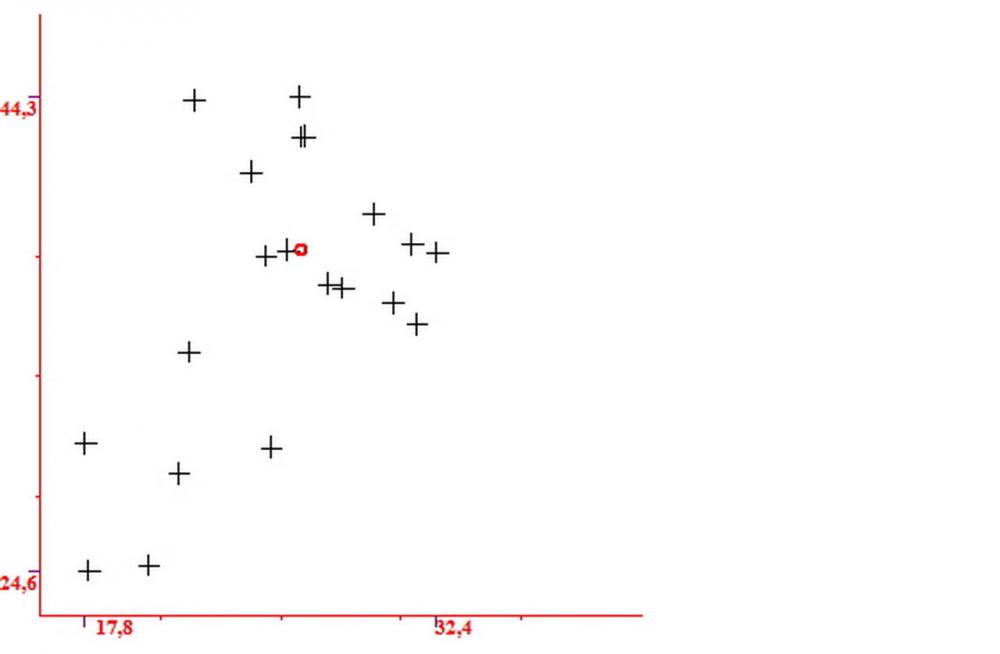
Pileipellis (24,5)25,1 - 36,0 - 44,0 (44,3) x (17,8) 18,2 -25,8- 31,5 (32,4) µm, Q. = 1,5-2,3, Q.m.= 1,88; Vol.m.= 13251,9 µm3, imeniderma con transizione in epitelio, composta da cellule più o meno pedicellate, ialine. Peli pileici 90-160 µm, plurisettati, sorgenti da una cellula inflata o più o meno piriforme, scarsi e limitati al margine.
Caulopellis (1,9)3,1 - 3,86 - 5,3 (6,0) µm, tipo cutis, formata da ife cilindriche, ialine.
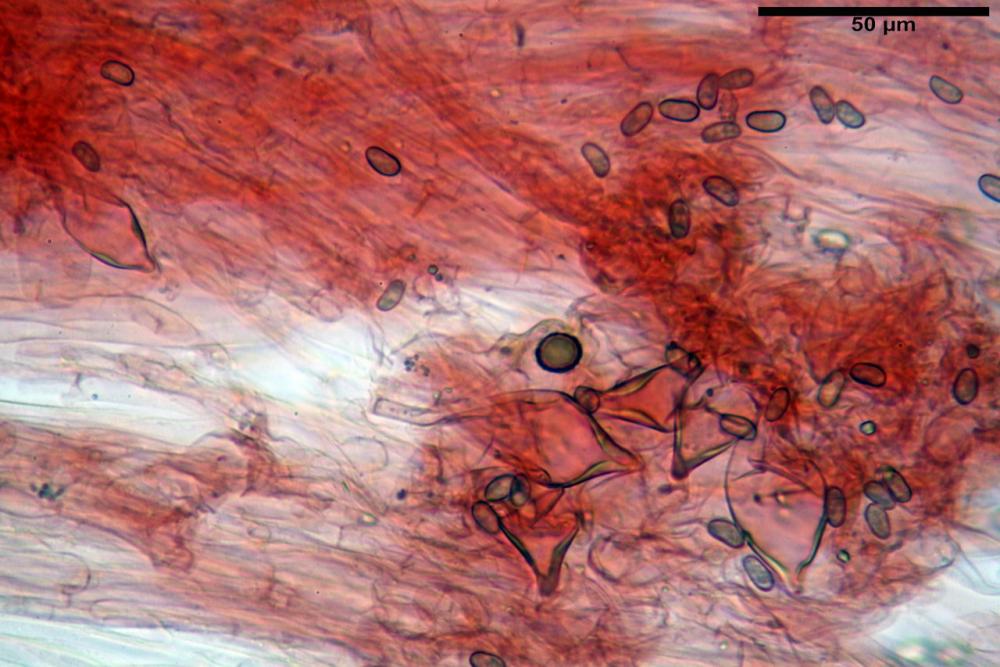
Caulocistidi metuloidi (32,5) 36,9 -44,1- 51,1 (57,6) x (12,0) 12,1 -15,9- 19,9 (20,9) µm, Vol.m.= 6053 µm3, simili ai cistidi imeniali, numerosissimi solo per i primi 2-3 mm, assenti più in basso.
Cauloleptocistidi (17,6) 19,4 -29,8- 43,8 (49,4) x (7,0) 7,7 -11,5- 18,1 (19,1) µm, Vol.m.= 2593 µm3, da clavati a subcilindrici, a parete sottile, ialini, presenti in radi mazzetti nella porzione superiore dello stipite, da rarefatti ad assenti più in basso.
Giunti a fibbia presenti in tutti i tessuti del basidioma.
Osservazioni
Nel lavoro sulla filogenetica molecolare delle Psathyrellaceae (Örstadius, Ryberg & Larsson, 2015), gli Autori convalidano formalmente il Genere Homophron e vi sistemano all’interno tre specie opportunamente ricombinate:
Homophron cernuum (Vahl) Örstadius & E. Larss., 2015, Homophron spadiceum (P. Kumm.) Örstadius & E. Larss., 2015 e Homophron camptopodum (Sacc.) Örstadius & E. Larss., 2015. Inoltre, confermando quanto già proposto in lavori precedenti (Örstadius, 2001 e Örstadius & Knudsen, 2008), riportano Psathyrella sarcocephala (Fr.) Singer 1951 e P. variata A.H. Sm. 1972 come sinonimi della nuova combinazione Homophron spadiceum.
Tra le specie psathyrelloidi Homophron spadiceum fa parte di un ristretto gruppo caratterizzato da cistidi imeniali metuloidi e muricati, cioè a parete relativamente spessa e con presenza di cristalli di ossalato di calcio.
Da parte nostra, e per la relativa poca esperienza su questa specie “solo tre raccolte studiate”, possiamo dire che abbiamo riscontrato caratteristiche piuttosto dissimili sia dal lato macroscopico che da quello microscopico.
Sebbene si accetti facilmente e si sorvoli spesso e volentieri sulle possibili differenze macroscopiche di “forme, colori, odori e sapori”, che una specie assume di volta in volta, adducendo a vari fattori causali e casuali, per es. temperatura, umidità ecc., non altrettanto facilmente si tende ad accettare queste differenze quando esse riguardano le caratteristiche microanatomiche di “basidi, cistidi, spore ecc.”, mentre sarebbe da rimanere più elastici anche con questi caratteri “intimi”, i quali, secondo il mio modesto punto di vista, sono suscettibili a modifiche, né più né meno, quanto quelli che si possono notare macroscopicamente.
Nelle tre raccolte studiate abbiamo riscontrato sostanziali differenze nelle misure dei cistidi metuloidi imeniali e nella loro forma. Nella quantità di cheiloleptocistidi, e/o la loro presenza/assenza. Nella presenza/assenza di caulocistidi metuloidi e/o la quantità e disposizione dei cauloleptocistidi. La presenza/assenza e/o abbondanza di muricazioni agli apici degli stessi. Le uniche identicità “se così si possono definire”, le abbiamo riscontrate nella muricazione dei pleurocistidi e nelle spore, simili sia nelle misure che nelle forme e nelle colorazioni.
Consultando la bibliografia in mio possesso ho potuto constatare come la specie allo studio, microscopicamente, sia molto simile, nelle forme e misure, a quella che veniva indicata e descritta come P. variata; inoltre alcune raccolte, non holotipiche, sub nomen P. variata sono state anche sequenziate e sono risultate conspecifiche con H. spadiceum. Non ci resta dunque che prendere atto della sinonimia sopra accennata. A tal proposito mi viene in mente un detto, che si usa spesso in Micologia, che recita così. -I funghi non sono francobolli-. E, aggiungerei io, sia macro che microscopicamente.
A parte le battute, malgrado attraverso queste mie osservazioni possa apparire che H. spadiceum possiede un intervallo differenziale molto esteso, dove alcune caratteristiche anatomiche possono variare ampiamente, vado ad elencare gli elementi stabili per una sicura e univoca determinazione di questo taxon:
- pleurocistidi muricati e fusoidi con apice acuto;
- spore di dimensioni medie (6,5-11 x 4-5,5 µm), pallide, con poro assente o indistinto.
Per ulteriori osservazioni sulle specie gravitanti attorno a H. spadiceum, si rimanda a questo link: http://goo.gl/wZ85cA
Ringraziamenti
Un sentito ringraziamento va all’amico Pietro voto per la revisione dell’Articolo ed a Mocnik Jože per la raccolta e le foto in Habitat.
Dati di raccolta e Habitat
N. scheda: 5594. Data di ritrovamento: 30/09/2015. Località: Kovcice. Comune: Hrpelje (SLO). Coordinate geografiche: 45°33' 47.35"N 14°04'24.81"E. Altezza s.l.m.: 558. Habitat: Bosco di latifoglia mista, su ceppaia marcescente di Alnus glutinosa. Determinatore: Zugna Marino, Mocnik Jože. Legit: Mocnik Jože.
Data inserimento in erbario: 13/10/2015.
Materiali e Metodi
Lo studio è stato compiuto su materiale fresco. I preparati sono stati osservati usando come mezzo di governo H2O; ove necessario, si è adoperato Rosso Congo Ammoniacale 2 % per meglio evidenziare le differenti tipologie cellulari. Le misure microscopiche sono state eseguite con il software di calcolo Mycométre gentilmente fornitoci dal Sig. Georges Fannechère e prelevabile a questo indirizzo http://mycolim.free.fr. Le misure sporali si riferiscono a 100 unità, effettuate in proiezione lateralesenza tener conto dell’apiculo o, nel caso di spore inequilaterali, anche in proiezione frontale, ottenute da sporata o prelevate da altre zone non imeniali, scartando spore ancora evidentemente immature, per le rimanenti tipologie cellulari si sono eseguite un minimo di venti misure a tipologia. Le misure sottolineate si riferiscono alle medie. Per la terminologia riguardante la tipologia delle caratteristiche macro e microscopiche, si fa riferimento a (Vellinga EC. 1988). Per quanto riguarda la nomenclatura aggiornata ed i nomi degli Autori, abbiamo fatto riferimento a http://www.indexfungorum.org e http://www.mycobank.org.
Le foto in habitat sono state eseguite con l’ausilio di una fotocamera COOLPIX P7800 e obiettivo, in studio, con fotocamera Reflex EOS 5D Mark II, obiettivo Canon EF 100mm f/2.8 Macro USM. Per le osservazioni dei caratteri macroscopici, in alcuni casi, ci siamo valsi di uno stereo microscopio Optech trinoculare e del supporto di foto macro ad alta risoluzione. Le foto concernenti la microscopia, sono state eseguite con l’ausilio di una fotocamera Reflex EOS 50D, posto sul terzo occhio di un microscopio biologico Optech Biostar B5, supportato da ottiche Plan-APO, illuminazione alogena 12V-50 W a luce riflessa, con regolatore d'intensità. Le collezioni d’erbario sono conservate nell'erbario A.M.B. Gruppo di Muggia e del Carso.
Bibliografia consultata
Arnolds E. (2003) Rare and interesting species of Psathyrella. Fungi Non Delineati 26:1–76.
Fouchier F., 1995 – Le Genre Psathyrella – Flore des espécies europeénne et méditerranéennes.
Hausknecht A., I. Krisai-Greilhuber (1997): Some rare Agaricales with brown or darkerspores. Fungi Non Delineati Pars II. Edizioni Candusso 1997.
Kits van Waveren, E. (1985). The Dutch, French and British species of Psathyrella. Persoonia Supplement 2.
Örstadius L. (2001): Psathyrella spadicea - taxonomy and nomenclature. Windahlia 24: 19–24.
Örstadius L. & Knudsen H. in Knudsen H. & Vesterholt J. (2008): Funga Nordica: 586-623 Ed. Knudsen & Vesterholt.
Örstadius L., Ryberg M. & Larsson E. (2015): Molecular phylogenetics and taxonomy in Psathyrellaceae (Agaricales) with focus on psathyrelloid species: introduction of three new genera and 18 new species. Mycol Progress (2015) 14: 25.
Vašutova M. (2008): Taxonomic studies on Psathyrella sect. Spadiceae. – Czech. Mycol. 60 (2): 137–171.
Vašutova M., Antonin V. and Urban A. (2008): Phylogenetic studies in Psathyrella focusing on sections Pennatae and Spadiceae– new evidence for the paraphyly of the genus. – Mycol. Res. 112 (10): 1153–1164.
Bibliografia citata
Vellinga EC. 1988. Glossary. In C Bas, Th W Kuyper, ME Noordeloos, EC Vellinga (eds.). Flora Agaricina Neerlandica 1: 54-64. A.A. Balkema. Rotterdam.
Dal Web
Forum di Micologia del Gruppo di Muggia e del Carso: http://www.ambmuggia...ea/?hl=spadicea
http://www.indexfung...Names/Names.asp
http://www.mycobank.org/MycoTaxo.aspx
Photo by: Mocnik Jože