Questa discussione è bloccata
Questa discussione è bloccata
Amanita malleata (Piane ex Bon) Contu, Bollettino dell'A.M.E.R. 3 (nos 6-7): 43 (1986)
Basionimo: Amanitopsis malleata Piane ex Bon, Docums Mycol. 12(no. 48): 33 (1983) [1982]
Sistematica: Amanitaceae, Agaricales, Agaricomycetidae, Agaricomycetes, Basidiomycota, Fungi
Quadro macroscopico della raccolta
Pileo 50-80 mm, non molto carnoso, dapprima globoso quindi semi convesso, infine disteso basso umbone e leggera depressione discale.
Cuticola apparentemente secca, untuosetta dopo umidificazione, totalmente, o quasi, separabile.
Allinizio di color ocra molto chiaro, poi con toni ocracei più sostenuti fino a ocra-brunastri al centro, impallidente verso il margine, orlo evidentemente striato per 1/5° di raggio. In gioventù decorata da lembi velari biancastri.
Lamelle mediamente fitte, larghe, spesse ma fragili, da rettilinee a leggermente ventricose, libere, biancastre nel giovane, crema chiaro con lieve riflesso rosa nelladulto (in exsiccata un esemplare ha manifestato tonalità lamellari fortemente rosate, infine sparite nellarco di un paio di giorni), filo concolore, intero o finemente fioccoso, lamellule presenti, tronche.
Stipite 100-140 x 10-15 mm, a lungo pieno poi farcito, quindi cavo, fragile, da cilindrico a svasato in alto e progressivamente allargato verso la base, non bulboso, biancastro allinizio quindi ocraceo, ornato da piccole squamette ascendenti e sottili fibrille nella parte bassa, bianco-ocraceo e finemente zigrinato in alto.
Volva bianca o solo occasionalmente ornata da piccole pustole ocracee, ocracea o appena imbrunente nella parte interna, da mediamente membranosa a bambagiosa, facilmente lacerabile.
Carne soda ma poco consistente, al taglio bianca nella polpa e nel midollo.
Odore nullo.
Sapore gradevole.
Reazioni chimico-cromatiche non effettuate.
Sporata in massa bianca.
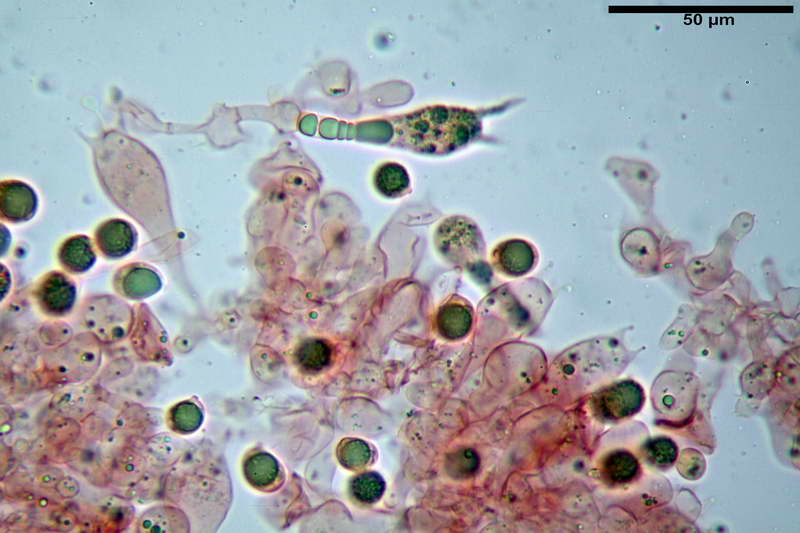
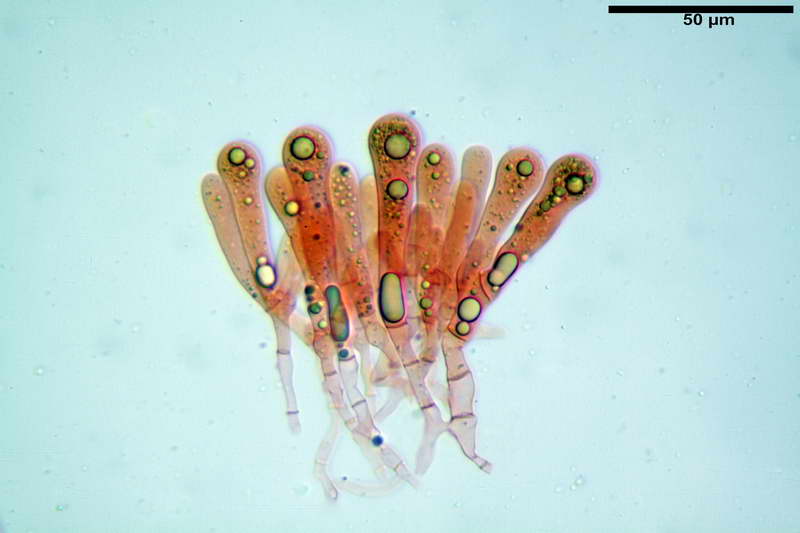
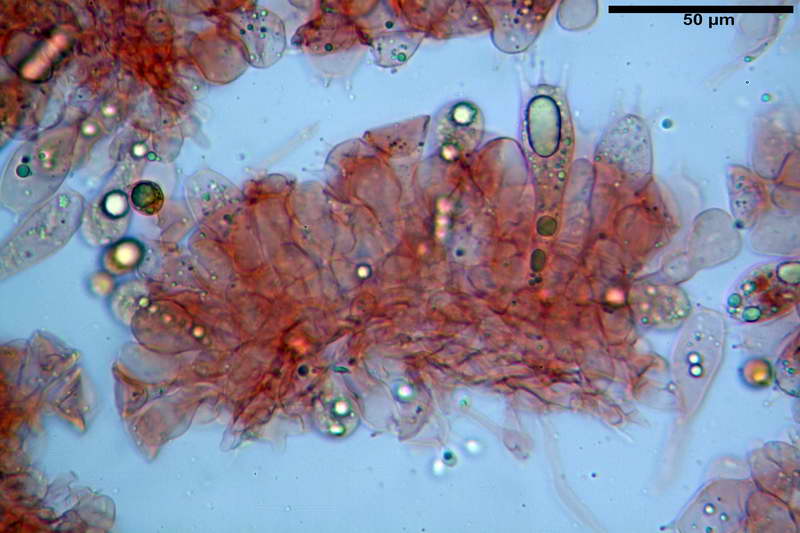
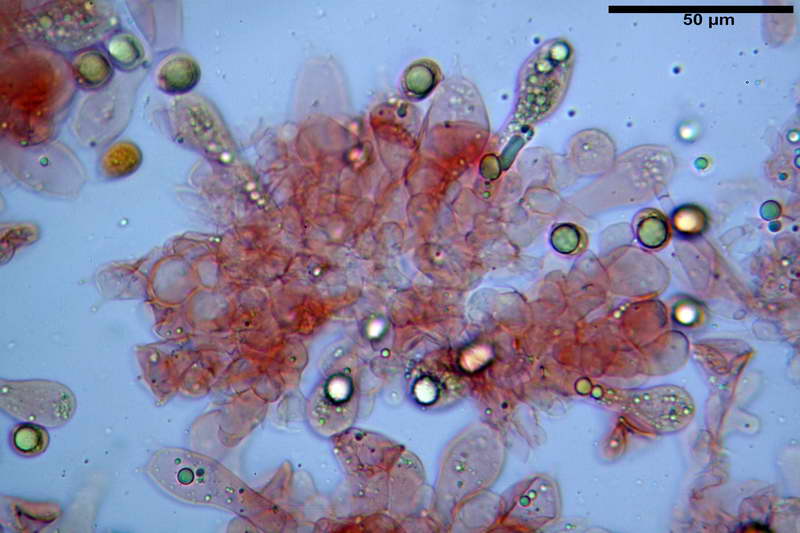
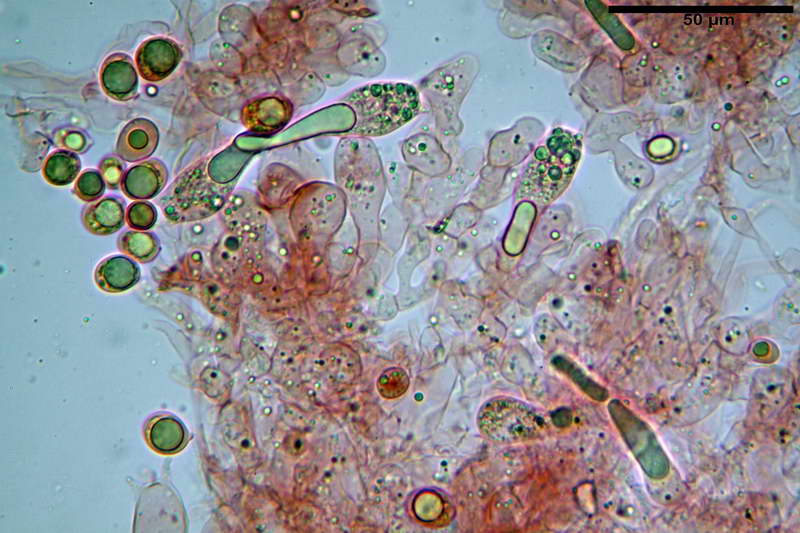
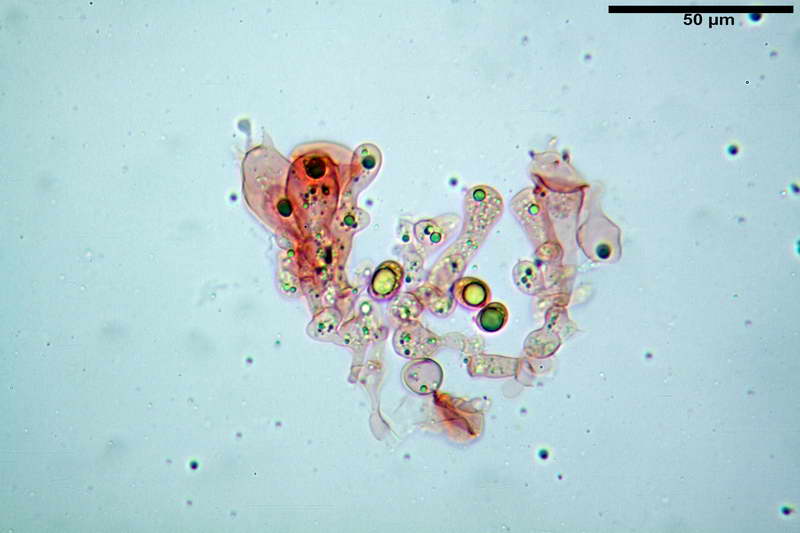
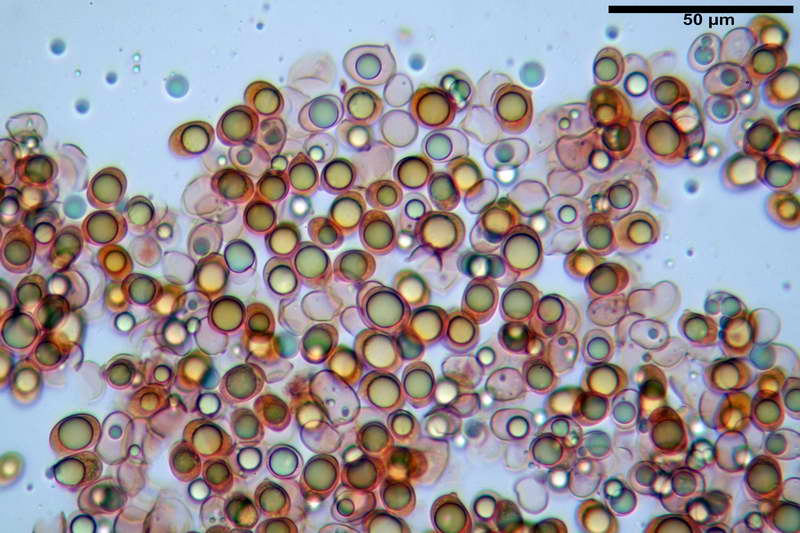
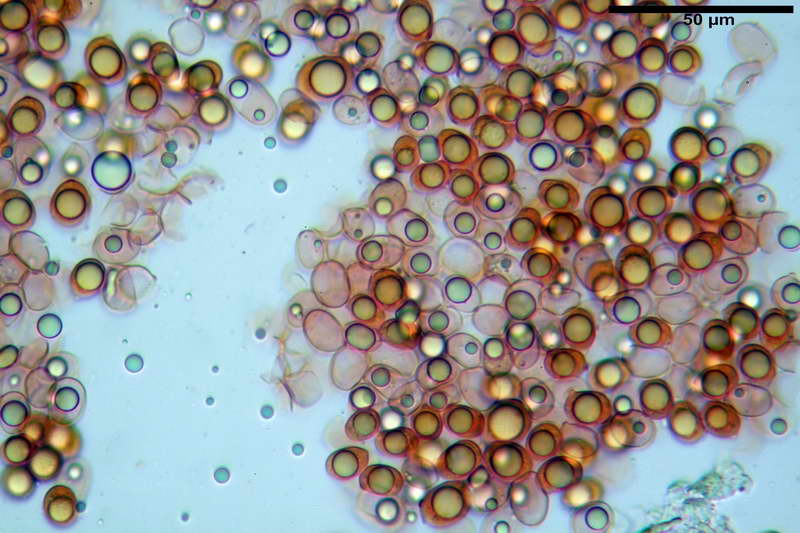
Quadro microscopico della raccolta
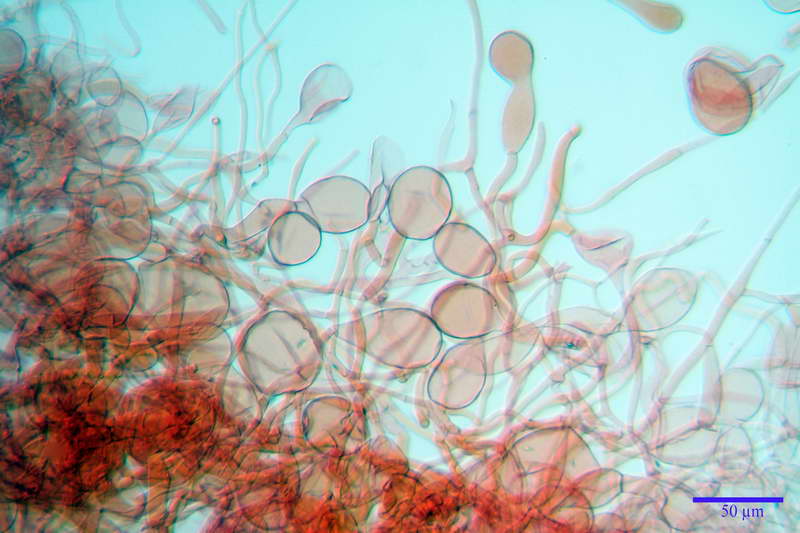
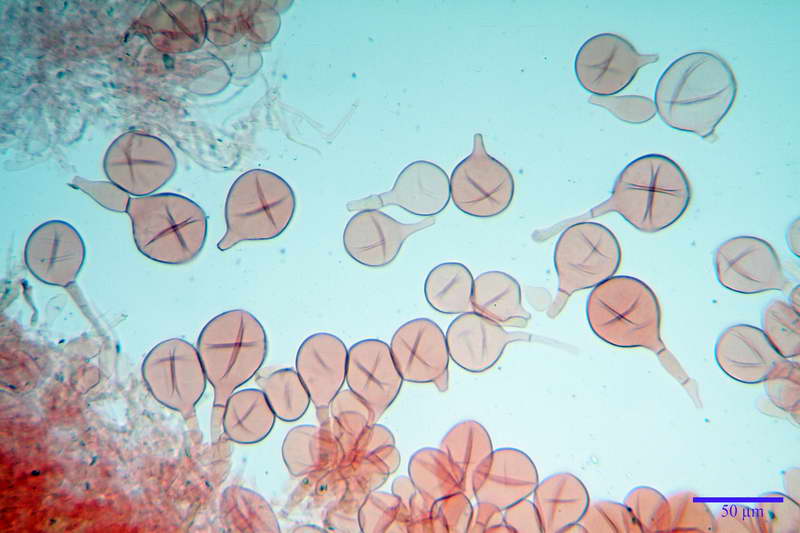
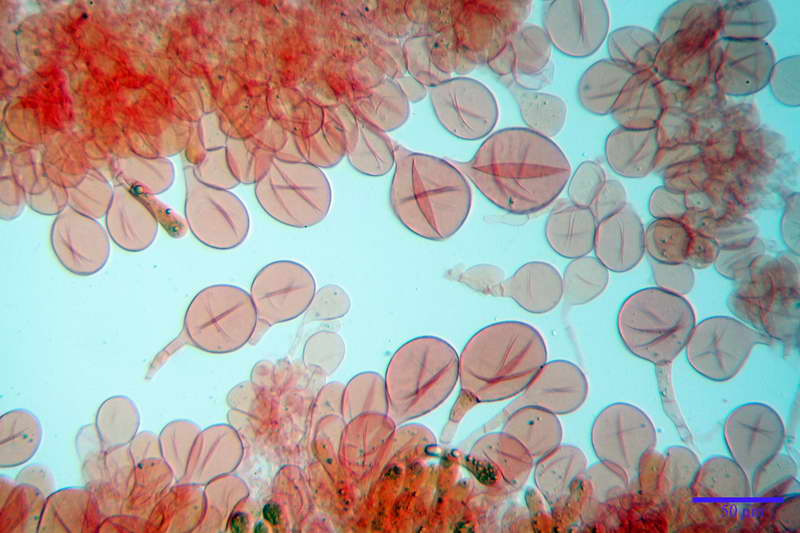
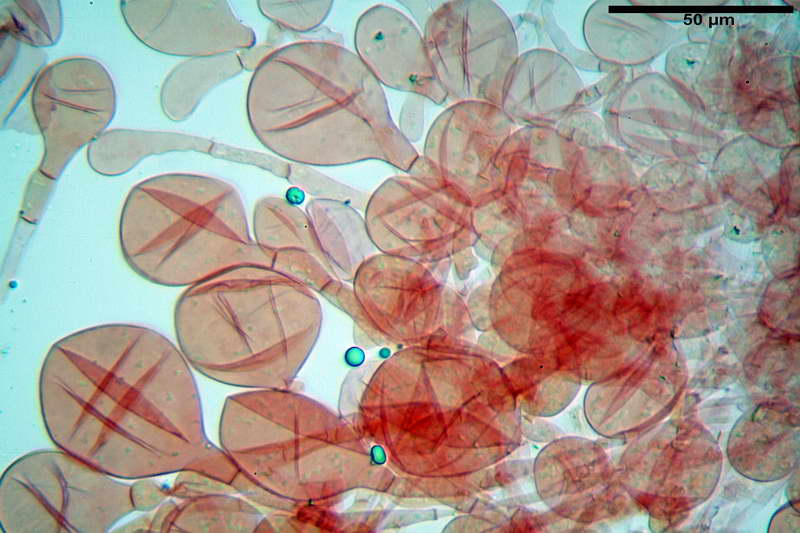
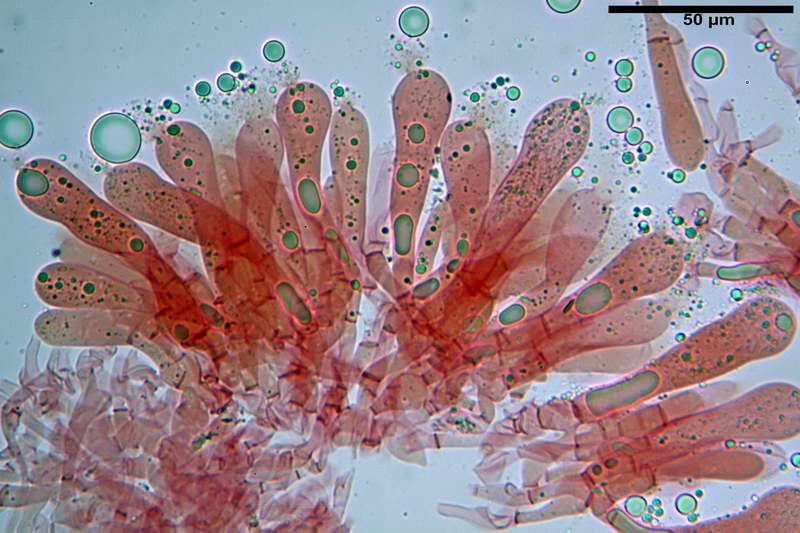
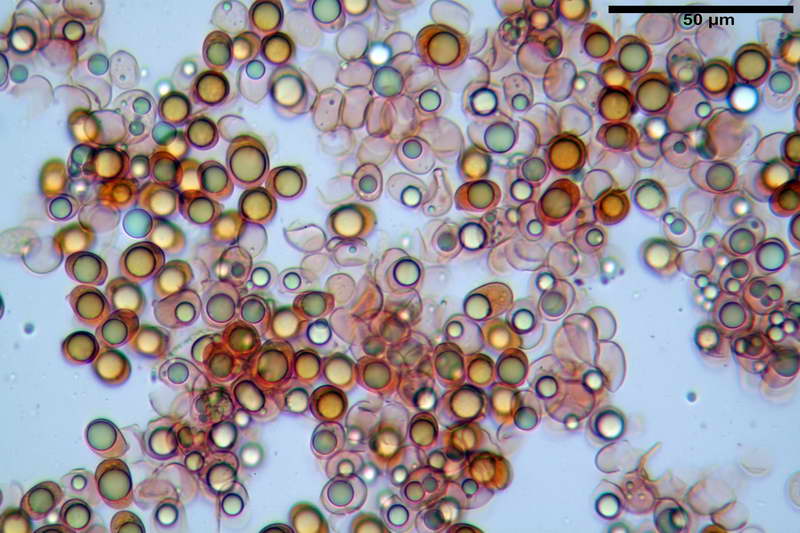
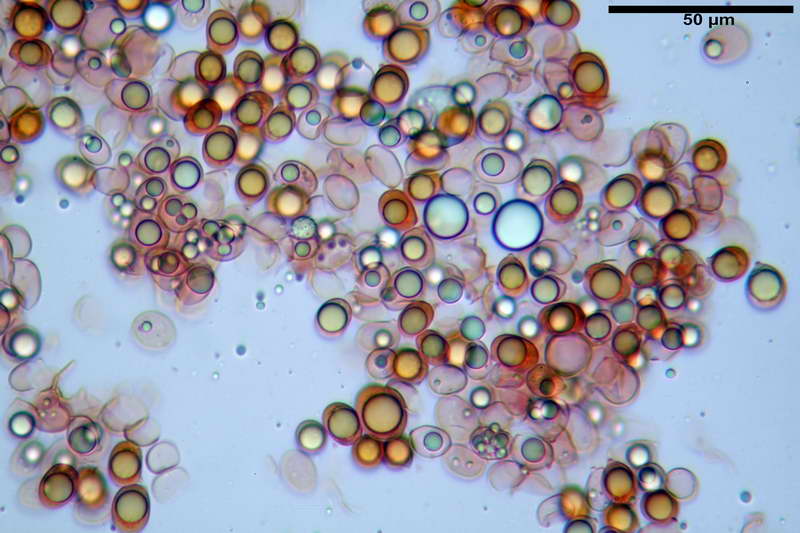
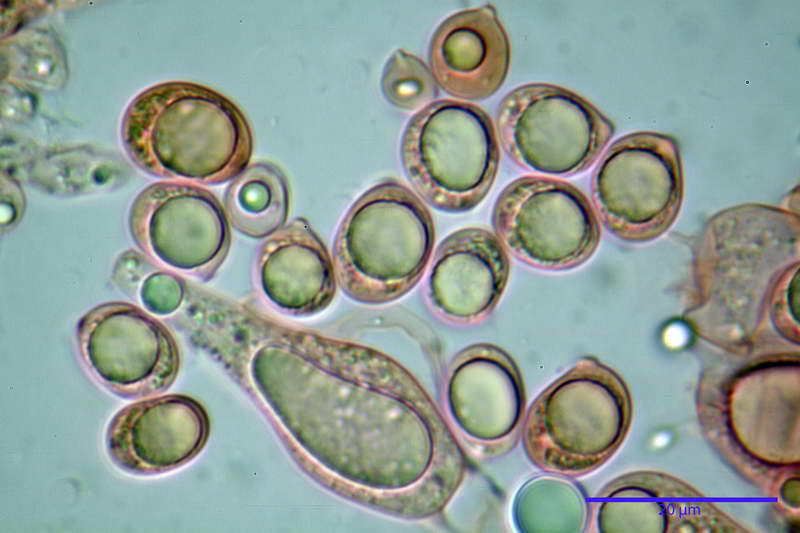
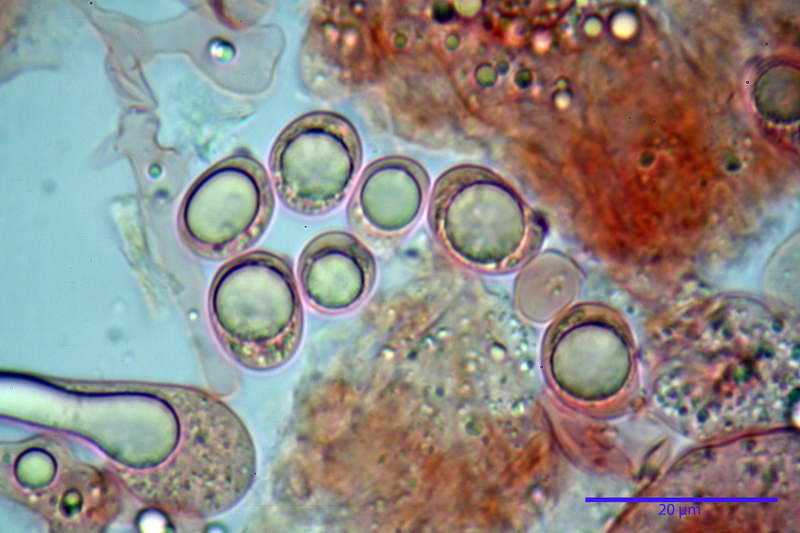
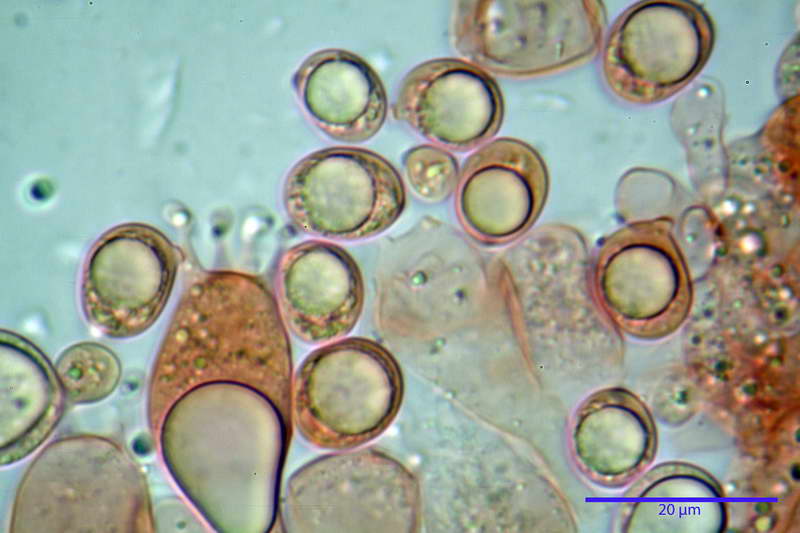
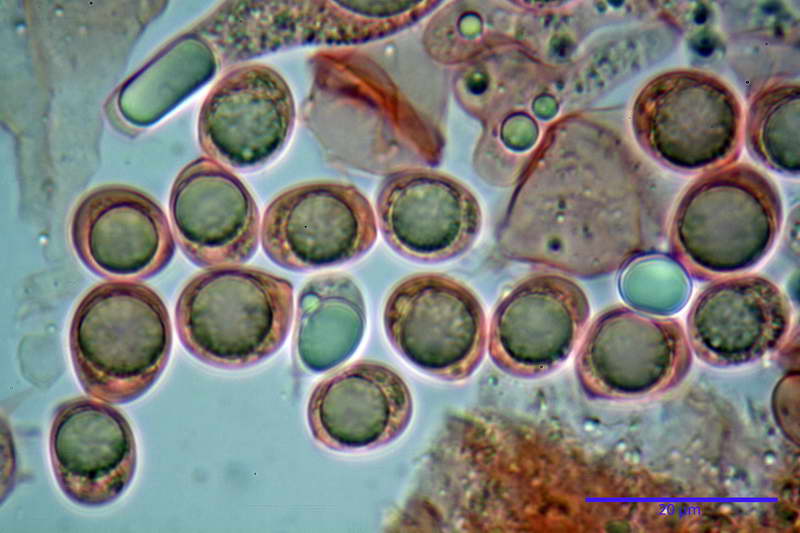
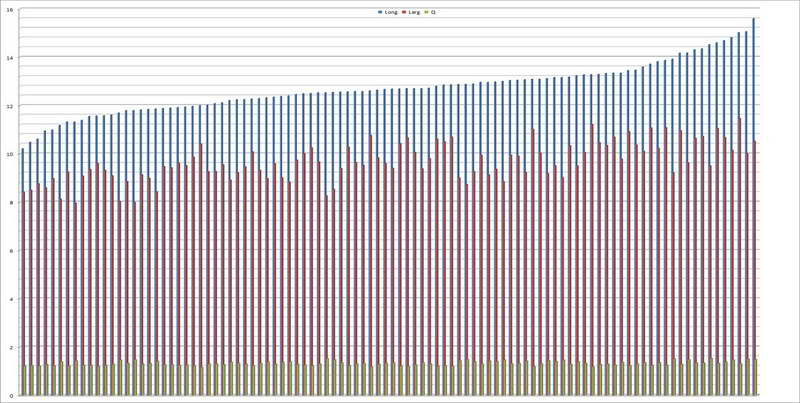
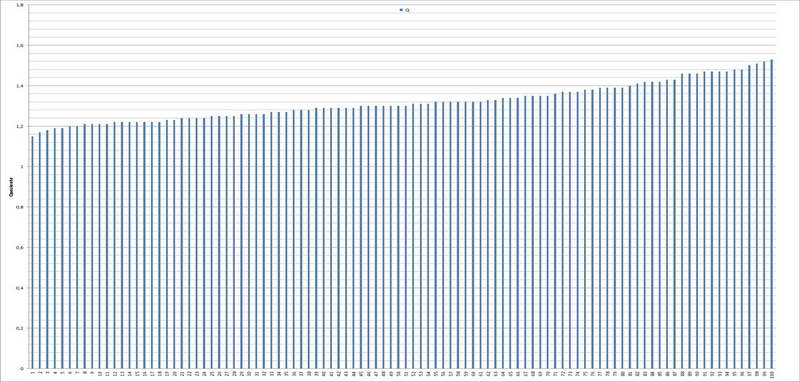
Spore (10,21) 11,55-14,17 (15,60) x (7,96) 8,60-10,71 (11,46) µm, in media 12,71 x 9,66 µm, Q. = (1,15) 1,21-1,46 (1,53), Q.m. = 1,32, Vol. = (375) 449-847 (1032) µm³ , Vol.m = 630 µm³, non amiloidi, ialine.
In proiezione laterale da largamente ellissoidi a ellissoidi; subglobose in proiezione ventrale, con una grande goccia oleosa centrale, apiculo prominente occasionalmente si sono notate spore leggermente lacrimoidi oppure subpiriformi.
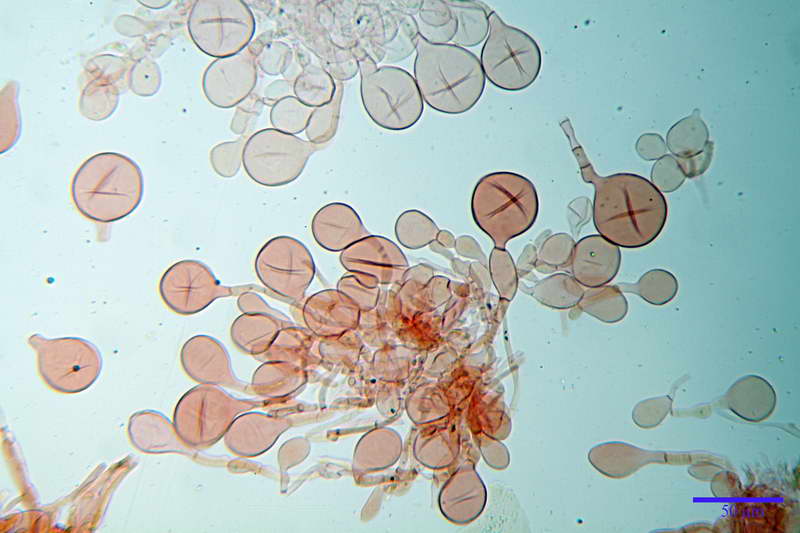
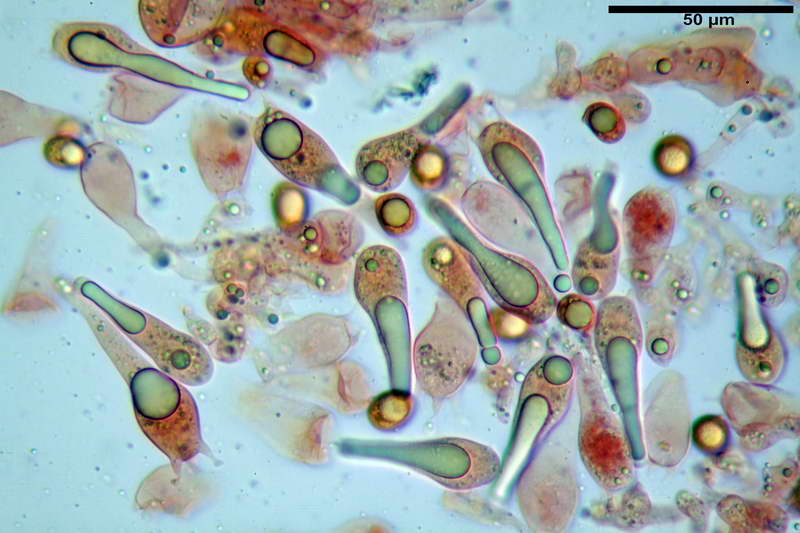
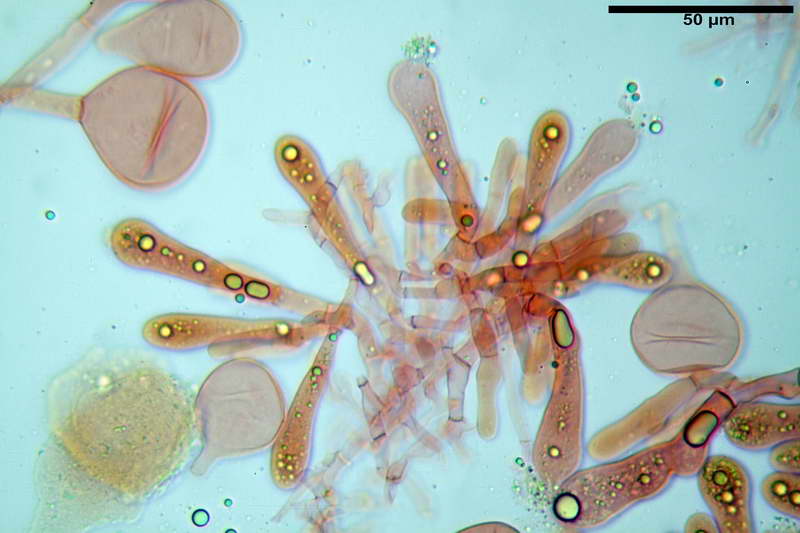
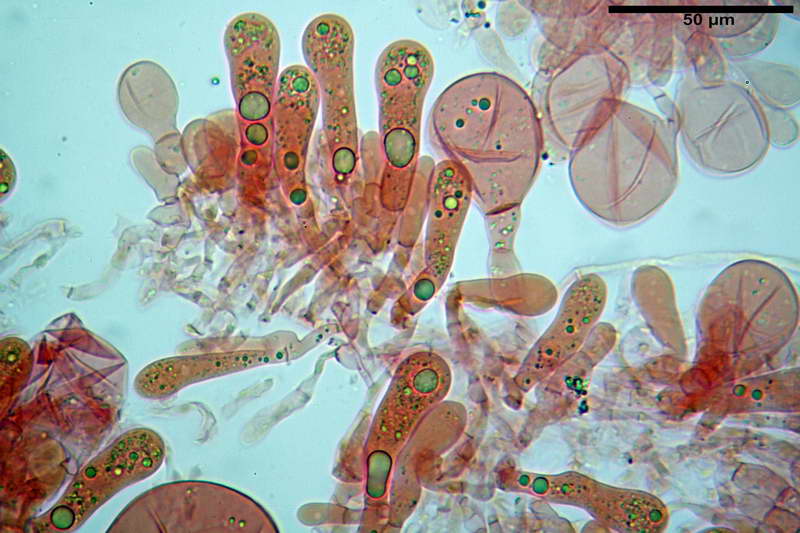
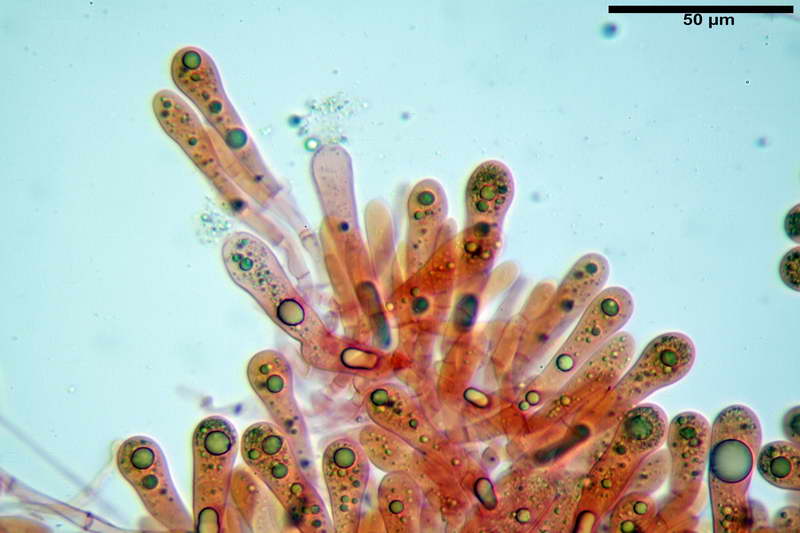
Basidi 46,2-66,4 x 12,9-17,4 µm, in media 53,43 x 14,92 µm, Q.m. = 3,58, Vol.m = 6330 µm³, in maggioranza tetrasporici ma anche diversi bisporici, claviformi, mediamente lunghi.
Albero subimeniale, in gioventù, sostanzialmente filamentoso, formato da una cellula ifale probasidiale di forma subcilindrica con Ø di 3,2-6,1 µm, Ø medio di 4,7 µm, seguita da una serie di cellule ifali, cilindriche, multi settate, con Ø di 2,0-4,0 µm, Ø medio di 3,3 µm, confluenti nella trama imeniale.
Negli esemplari maturi laspetto dellalbero imeniale tende a modificarsi notevolmente, assumendo un aspetto prismatico con le prime due, tre cellule, tendenti ad allargarsi fino a un Ø di 5,0-12,7 µm, con un Ø medio di 8,14 µm, mentre quelle sottostanti mantengono laspetto e la forma originaria.
Trama imeniale divergente.
Laticiferi non osservati.
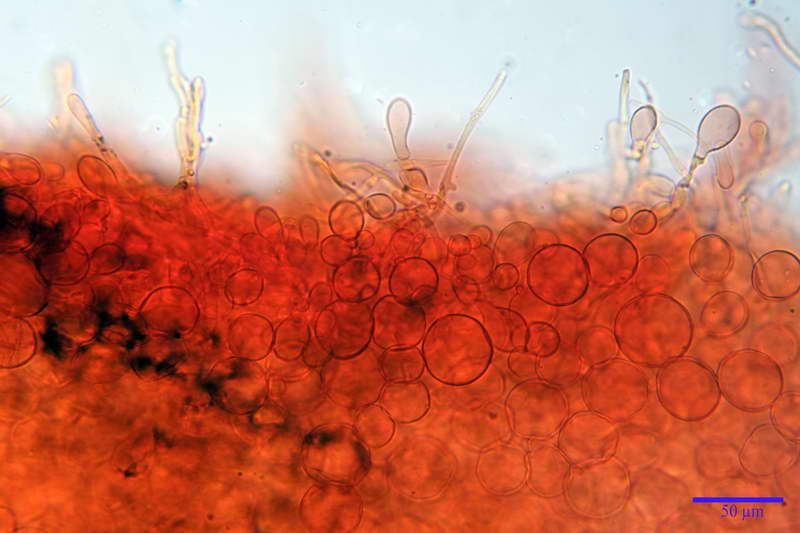
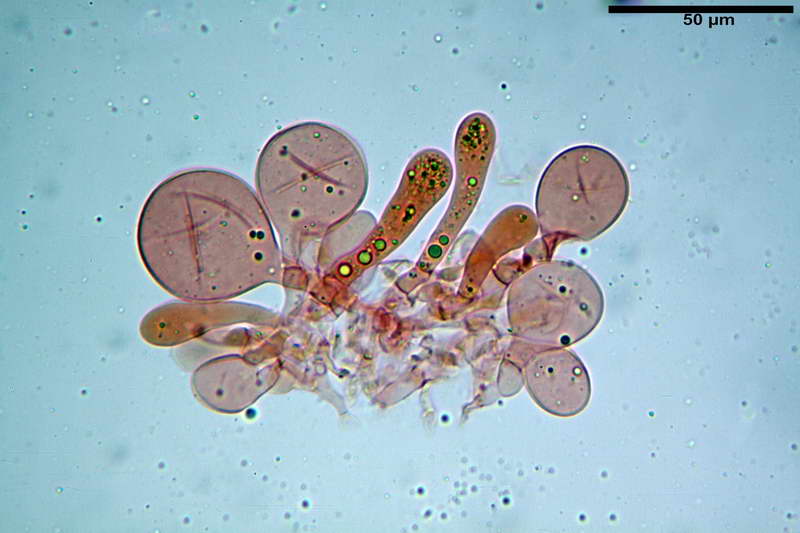
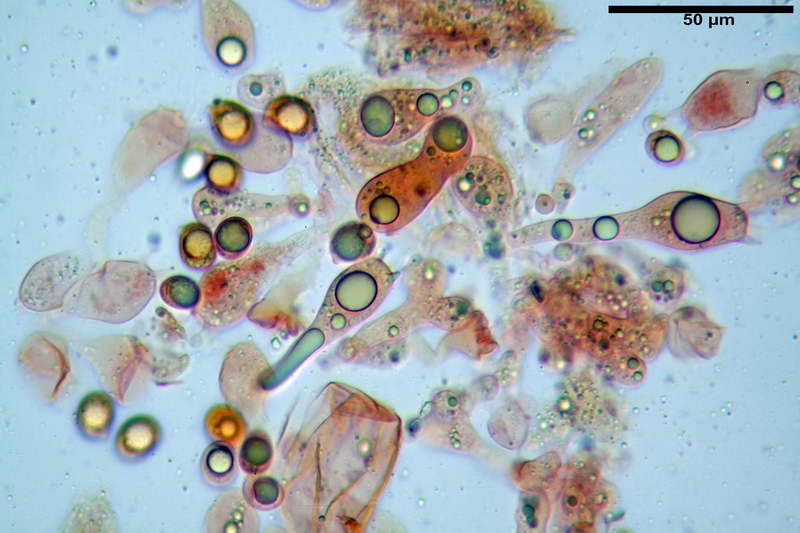
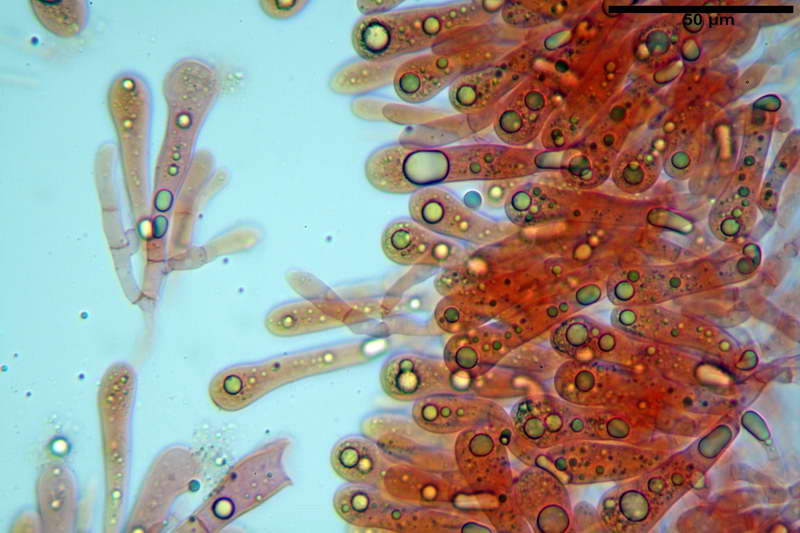
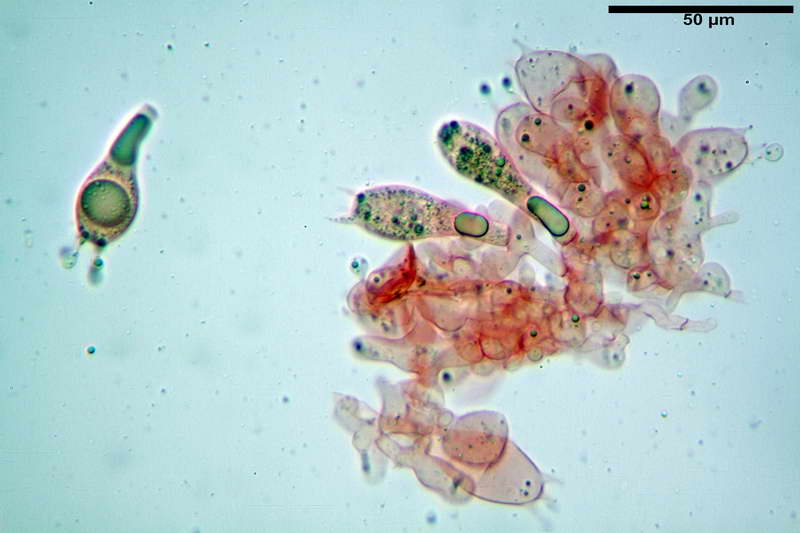
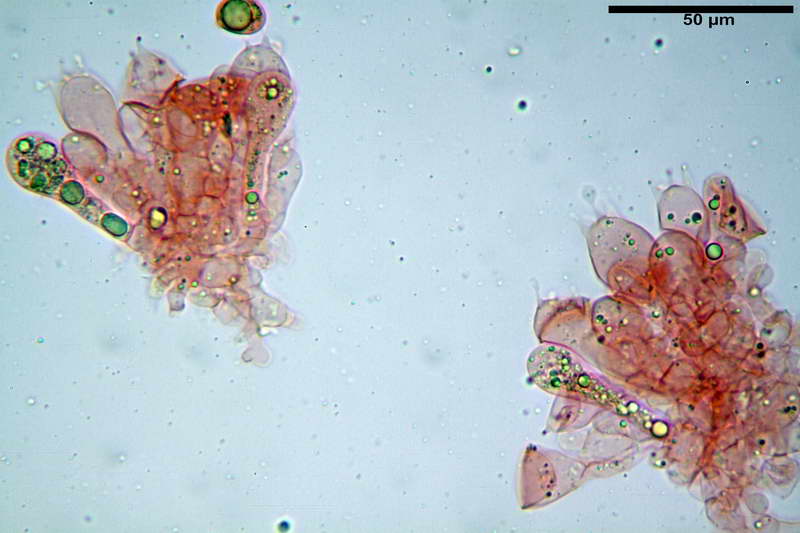
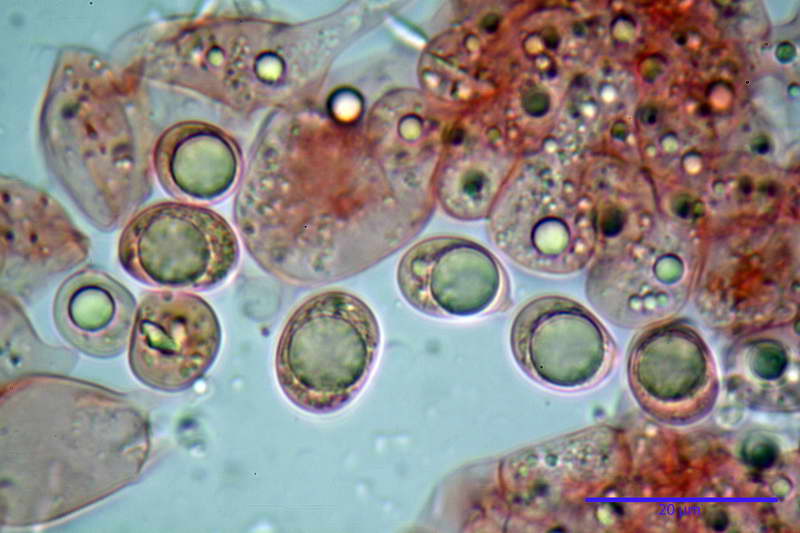
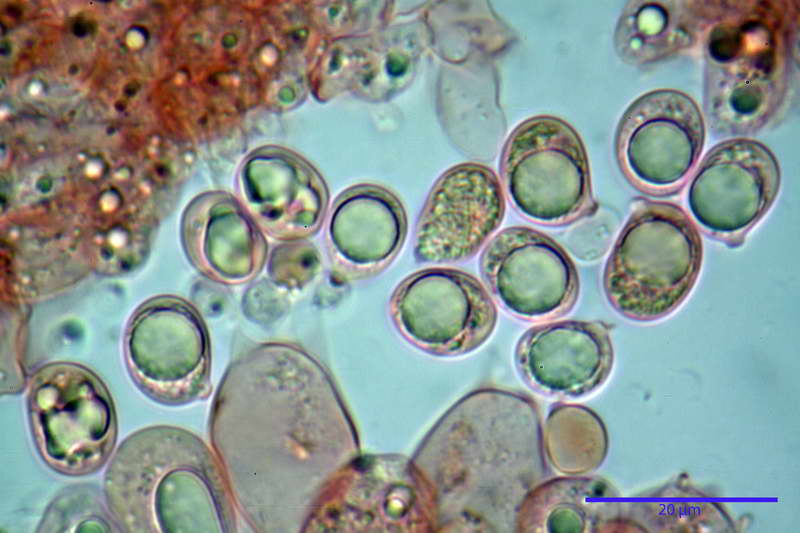
Cellule marginali (33,1) 35,0-54,1 (60,3) x (12,0) 27,0-41,7 (44,1) µm, in media 46,90 x 34,74 µm, Q.m. = 1,38, Vol.m = 31614 µm³, a parete sottile, ialine, nella stragrande maggioranza di forma sferopeduncolata, raramente clavate, spesso con uno, due o tre setti basali; negli esemplari giovani tappezzanti lintero filo lamellare, negli esemplari adulti da rade ad assenti.
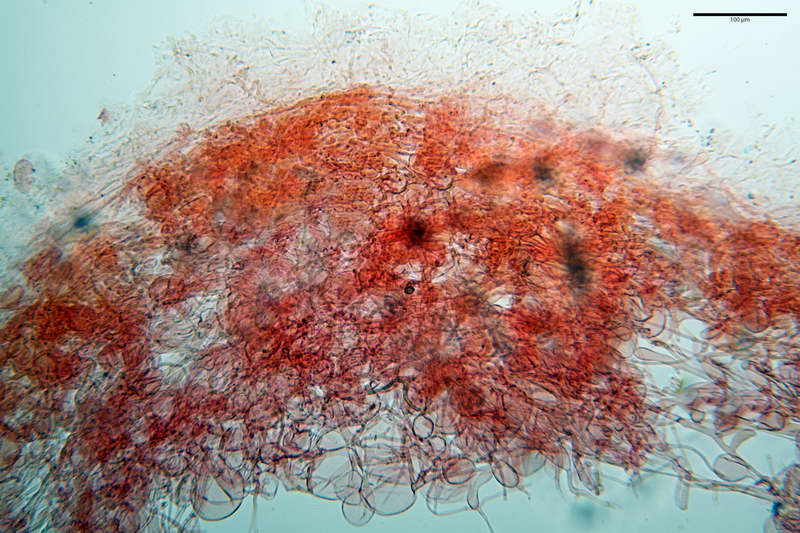
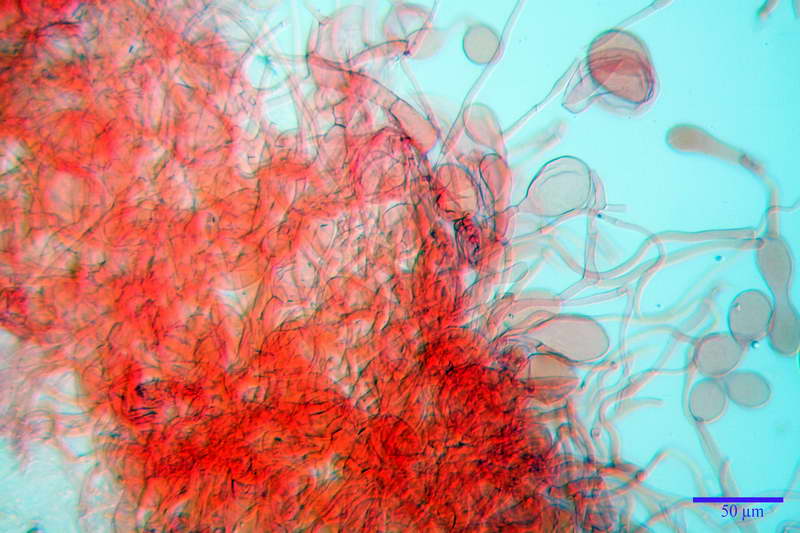
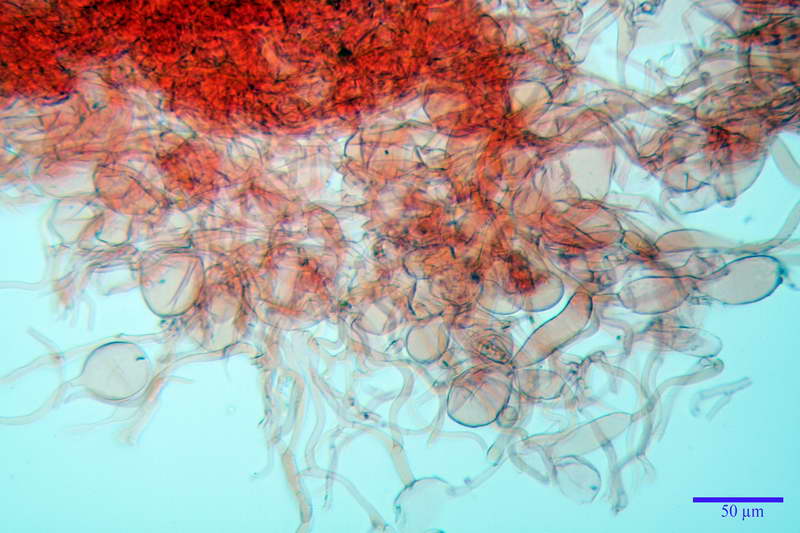
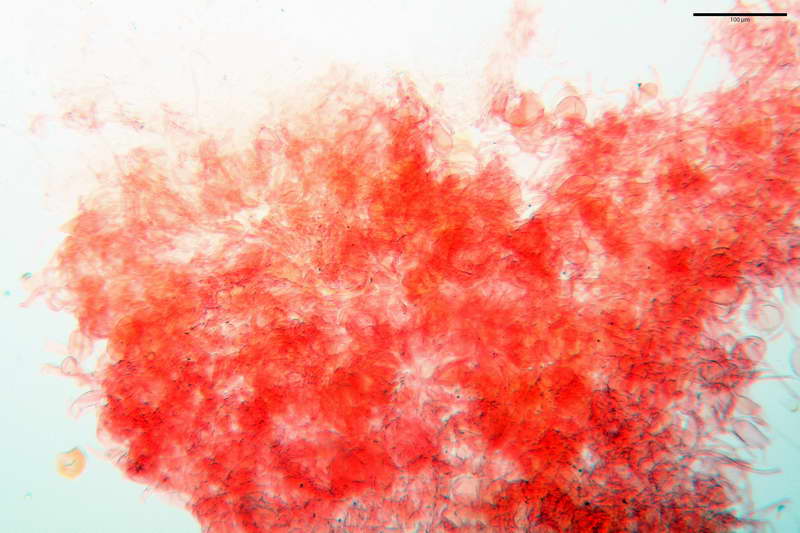
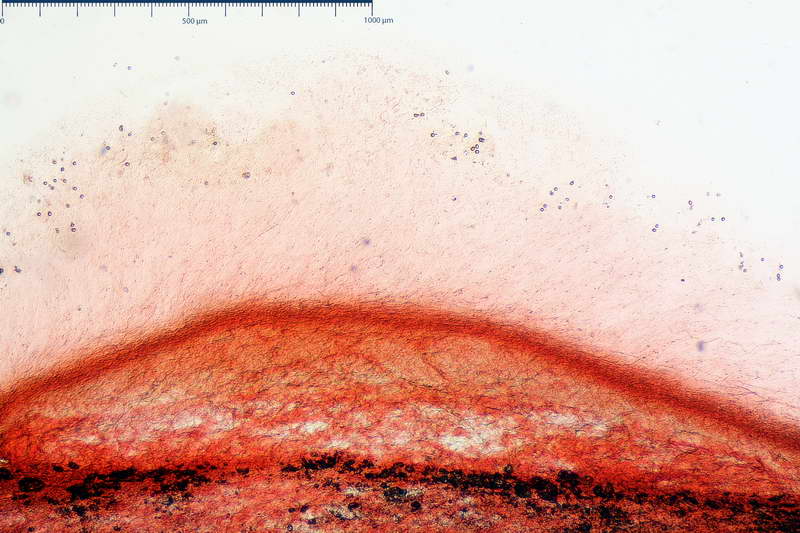
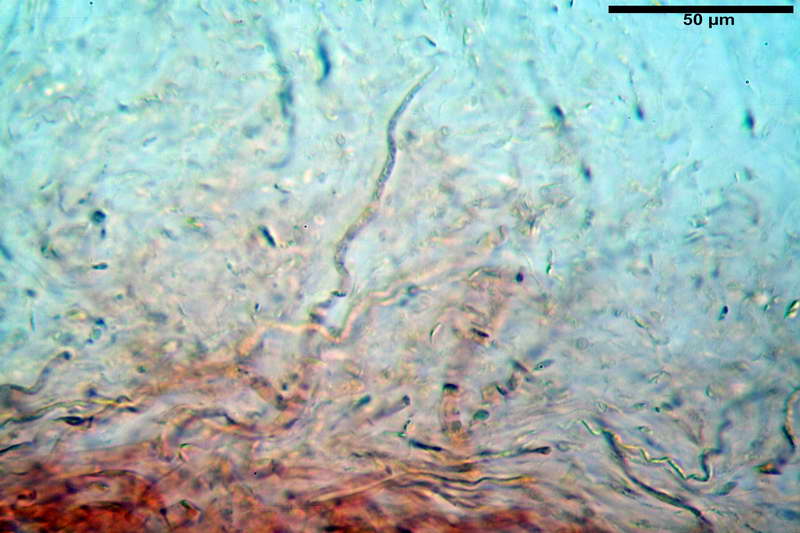
Velo generale prelevato dallorlo dalla volva.
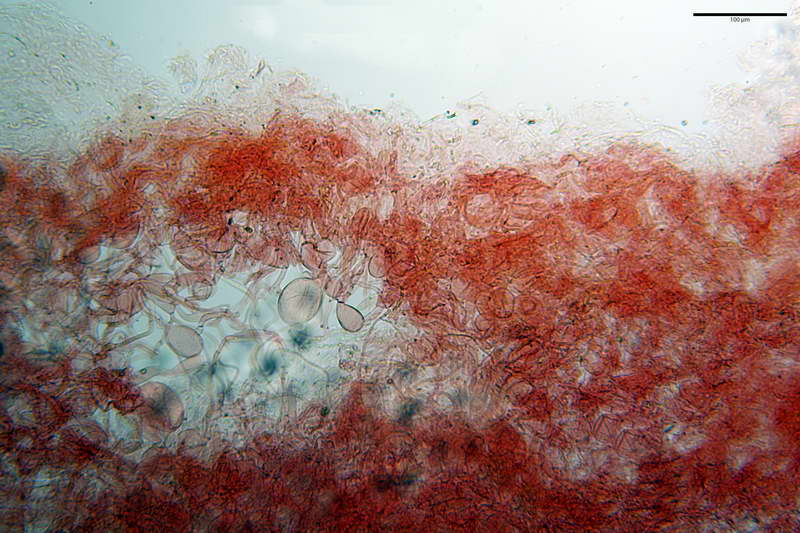
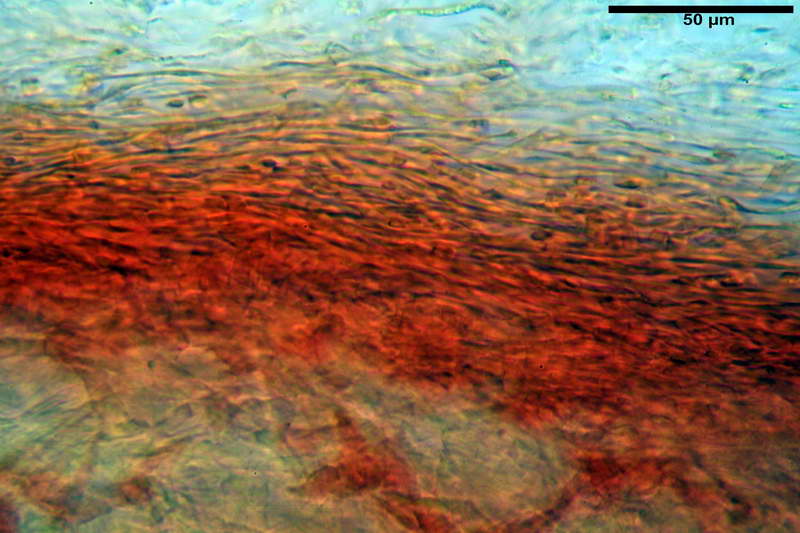
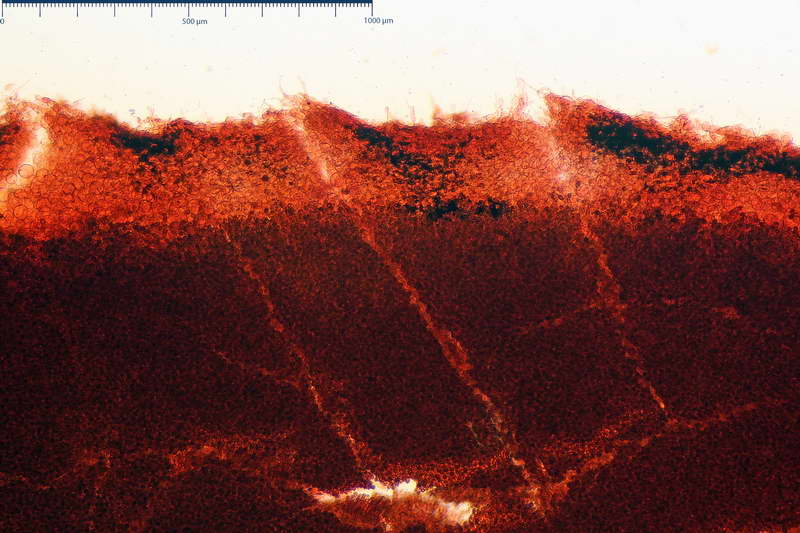
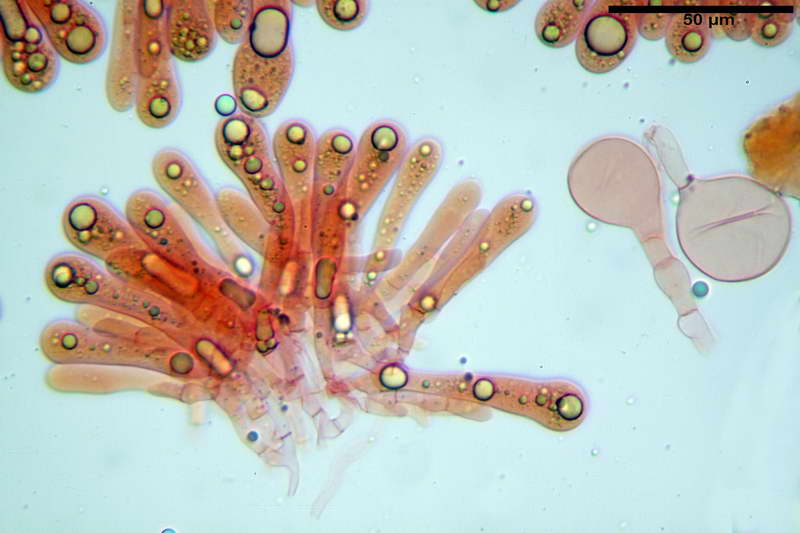
Parte esterna della volva in ixocutis spessa 67-160 µm, formata da ife gelatinizzate, congofile, ialine.
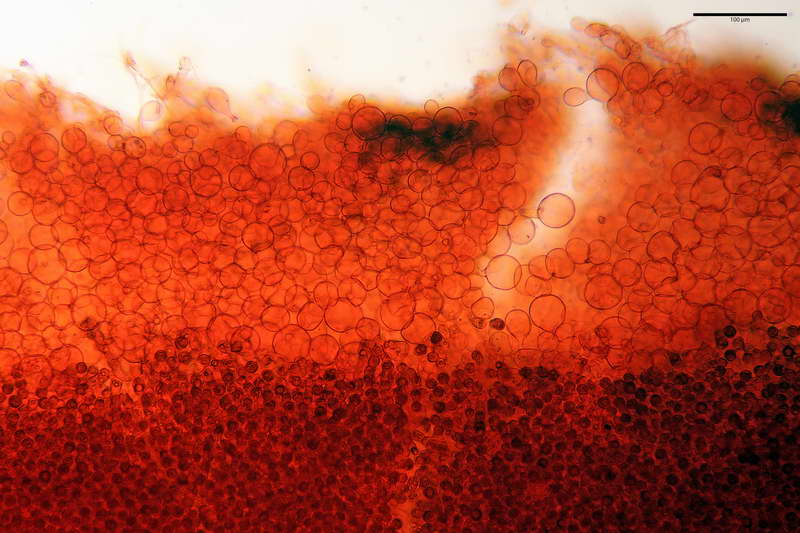
Lo strato più esterno, sostanzialmente ifale e composto da poco numerosi cilindrocidi e ife congofobe, spesso allincirca 20-45 µm, a cui segue uno strato centrale, di maggior spessore, misto, formato da un numero abbastanza cospicuo di sferociti e ife filamentose in egual misura, per terminare con un terzo strato, più interno, simile a quello esterno per conformazione e spessore.
Sferociti, cellule sferopeduncolate, largamente o strettamente clavate misuranti (28,5) 34,5-68,5 (95,0) x (18,5) 22,4-61,3 (74,5) µm, in media 49,6 x 36,5 µm, Q.m. = 1,43, Vol.m = 48053 µm³.
Cilindrocidi con Ø di 10,5-19,0 µm, Ø medio di 13,8 µm, a parete sottile e ialini.
Ife filamentose ramificate, settate, prive d incrostazione con Ø di 2,7-5,7 µm, Ø medio di 4,2 µm; terminali di pari spessore.
Laticiferi non osservati
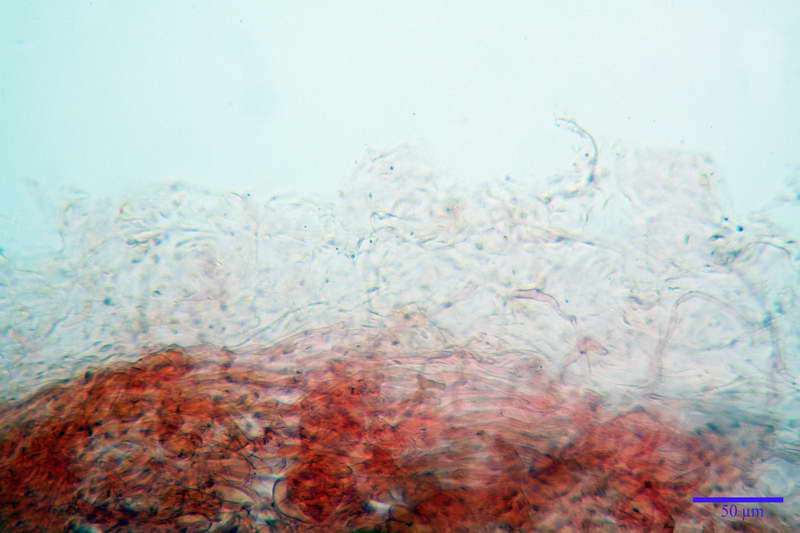
Rivestimento pileico in ixocutis spessa 350-500 µm, composta da gracili ife cilindriche, gelatinizzate, congofile, ialine, con Ø di 1- 4 µm.
Pileipellis tipo cutis, formata da ife cilindriche congofobe, coricate e debolmente intrecciate, con Ø 3-7 µm, presenza di pigmento vacuolare. Laticiferi presenti nella subpellis.
Giunti a fibbia non osservati in nessun tessuto del basidioma.
Osservazioni
A dire la verità, allinizio non eravamo certi che il taxon da noi ipotizzato fosse il più adatto.
Troppe, in passato, sono state le interpretazioni differenti, (soprattutto di spore e struttura del velo), che gli AA., hanno attribuito ad A. malleata. In effetti, cè chi la considera una specie a spore subglobose con Q.m.=1.0-1.1, chi, invece, una specie con spore da largamente ellissoidi a ellissoidi e con Q.m.=1.15-1.5.
Per quanto riguarda precedenti raccolte, da noi attribuite a questo taxon, il Q.m. è sempre stato quello di spore largamente ellissoidi e di seguito riportiamo i dati dei quozienti:
Ex.2365: Q. = 1,21-1,53, Q.m. = 1,37.
Ex.2640: Q. = 1,18-1,46, Q.m. = 1,32.
Ex.2812: Q. = 1,14-1,45, Q.m. = 1,29.
Ex.2818: Q. = 1,19-1,38, Q.m. = 1,28.
Basandoci sulle indicazioni della letteratura in nostro possesso (Contu 2001), abbiamo potuto appurare che A. malleata possiede spore largamente ellissoidi, misuranti 10-13,5 x 8-11 µm (Contu 2001).
Di seguito, tentiamo di formulare alcune brevi considerazioni .
La raccolta in oggetto ci pone di fronte un taxon con spore da largamente ellissoidi a ellissoidi, il che, sommato alle ulteriori caratteristiche macro e microscopiche, ben si adatta alla succitata specie (altrimenti dovremmo considerare la possibilità di una specie non ancora descritta).
Dal lato sistematico, stabilito che, la raccolta in oggetto possiede un velo generale ifale misto ad abbondanti sferociti e un Q.m. = 1,32, non dovremmo avere dubbi sul suo inserimento nella Sezione Vaginatae, sottosezione Ovigerae, serie biovigera (Contu 2001).
La serie è comprensiva di altri tre ulteriori taxa:
A. biovigera possiede un pileo di colore bruno-olivastro e predilige i boschi di conifere;
A. oblongispora possiede basidi con giunti a fibbia e predilige i terreni calcarei;
A. dryophila pileo bronzeo-aranciato a fulvo-nocciola e gambo con screziature concolori anchessa con preferenza per i terreni calcarei.
Ulteriori note in merito allo studio .
Per la misura delle spore abbiamo preso in considerazione solo le spore con apiculo ben evidente e posto lateralmente, in modo da essere sicuri che la spora sia situata sia in proiezione laterale, tralasciando spore con apiculo posto in posizione centrale, oppure senza apiculo evidente.
Losservazione, riguardo allalbero subimeniale, di esemplari immaturi e maturi è stato eseguito di proposito per fare un esempio e le notevoli differenze si intuiscono già da subito, in quanto, oltre allevidente differenza strutturale, lalbero subimeniale immaturo evidenzia basidi totalmente privi di sterigmi e totale assenza di spore adiacenti, mentre lalbero subimeniale maturo evidenzia basidi con lunghi sterigmi e spore in abbondanza.
A tal riguardo, alcuni AA., considerano la struttura dellalbero subimeniale non significativa dal punto di vista tassonomico, in quanto estremamente incostante, data la sua estrema variabilità strutturale, dallo stadio immaturo a quello stabile, finale.
A mio avviso non cè da meravigliarsi del fatto che la struttura del subimenio si sia modificata, in quanto, si tratta semplicemente del grado naturale di sviluppo delle cellule che, maturando, aumentano e prendono forma finché arrivano ad avere il loro aspetto adulto.
Altrimenti, allo stesso modo, ci dovremmo meravigliare del fatto che i basidi immaturi siano privi di sterigmi, mentre a maturazione li possiedano; oppure che, in altri casi, delle spore immature non siano pigmentate e/o ornamentate, mente, a maturazione, assumano la pigmentazione e lornamentazione definitiva.
Quello che è certo e che, se lo si vuole prendere in considerazione come carattere tassonomico stabile, ci si deve basare, sempre e solo, su cellule completamente mature.
In fin dei conti quando mai, per misurare spore, basidi, cistidi e quantaltro, ci si è basati su cellule immature!
Spero che l esempio posto, possa essere una buona dimostrazione e far riflettere in merito.
Materiali e Metodi
Lo studio è stato compiuto sia su materiale fresco sia secco; quest'ultimo, reidratato in KOH 2%, in Ammoniaca 2% o in acqua secondo i casi.
I preparati sono stati osservati usando come mezzo di governo H2O; ove necessario, si è adoperato Rosso Congo Ammoniacale 2% per meglio evidenziare le differenti tipologie cellulari. Il reagente di Melzer è stato utilizzato per laccertamento di uneventuale amiloidia sporale.
Le misure microscopiche sono state eseguite con il software di calcolo Mycométre gentilmente fornitoci dal Sig. Georges Fannechère e prelevabile a questo indirizzo http://mycolim.free.fr.
Le misure sporali si riferiscono a 100 unità, per le rimanenti tipologie cellulari si sono eseguite un minimo di venti misure a tipologia.
Per le osservazioni dei caratteri macroscopici, in alcuni casi, ci siamo valsi di uno stereo microscopio Optech trinoculare e del supporto di foto macro ad alta risoluzione.
Le foto macro sono eseguite in habitat, con lausilio di una fotocamera Reflex EOS 50D + obiettivo Canon EF 100mm f/2.8 Macro USM.
Le foto concernenti la microscopia, sono state eseguite con il corpo macchina della medesima fotocamera, posto sul terzo occhio del trinoculare di un microscopio biologico Optech Biostar B5, supportato da ottiche Plan-APO, illuminazione alogena 12V-50 W a luce riflessa, con regolatore d'intensità.
Le collezioni derbario sono conservate nell'erbario A.M.B. Gruppo di Muggia e del Carso.
Dati relativi alla raccolta N. scheda: 4475. Data di ritrovamento: 18/08/2010. Località: SANTA BARBARA. Comune: MUGGIA (TS). Coordinate geografiche: 45°35'8.90"N 13°47'8.84"E. Altezza s.l.m.: 207. Habitat: Ai margini di un prato boschivo, in terreno argilloso con sottosuolo di arenaria, nei pressi di Quercus pubescens, Ostrya carpinifolia, Fraxinus ornus. Determinatore: Zugna M. Legit: Zugna M.
Bibliografia consultata
CONTU M., 2000: Chiave per la determinazione delle specie europee del genere Amanita sez. Vaginatae. Boll. Gr. Micol. Bresadola 43: 233-240.
CONTU M., 2001: A revised key to Amanita section Vaginatae (Fr.)Quél. in Europe. Field Mycology 4(4): 128-136.
FRAITURE A.,1993: Les Amanitopsis dEurope. Op. Bot. Belg. 5: 1-127.
KÜHNER R. & ROMAGNESI H. ,1953 : Flore Analitique des Champignons Superieurs. Paris.
TRAVERSO M. 1999: Il genere Amanita in Italia. Roma.